Проведение возбуждение по нервным волокнам
В основе развития процесса проведения возбуждения по нервным волокнам наряду с ПД участвуют пассивные электрические свойства мембраны нервных волокон, которые определяются на эквивалентной схеме нервного волокна, погруженного в проводящую среду (Рис):
Сопротивление мембраны (rm) рассчитывается через удельное сопротивление (Rm):
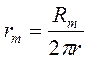 ,
,
где: Rm=1-100 кОм·см2, в перехватах Ранвье: 30-40 Ом·см2
Сопротивление аксоплазмы (ri) рассчитывается через удельное сопротивление (ri):
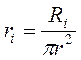 ,
,
где: Ri=40 Ом·см.
Емкость мембраны (cm) рассчитывается через удельную емкость мембраны (Сm):
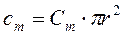 ,
,
где: Сm=1мкФ/см2, в перехватах Ранвье:3-4 мкф/см2, r –радиус нервного волокна, R,C –удельные и r,c- относительные значения сопротивления и емкости мембраны (m) и аксоплазмы (i) в рассчете на единицу длины или площади мембраны.
Возвращаясь к выше представленному выражению для расчета мембранного тока:
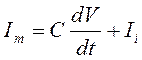
следует определить согласно эквивалентной электрической схеме, что мембранный ток аксиального направления будет равен согласно закону Кирхгофа:
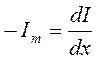 ,
,
В свою очередь согласно закону Ома ток, текущий через аксоплазму равен:
 , а итоговый:
, а итоговый: 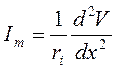 .
.
В окончательном виде:
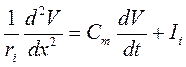 ;
;
 ;
; 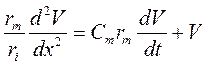 .
.
С учетом того, что
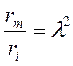 и
и 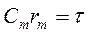 ,
,
окончательное выражение принимает вид:
 .
.
Решение этого уравнения требует ряд упрощений:
1. При аксиальном введении регистрирующего электрода мембранный ток становится равным нулю и тогда:
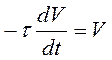 ,
,
решением этого уравнения является экспоненциальная зависимость:
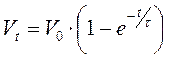 ,
,
где τ – постоянная времени, показывающая через сколько времени амплитуда мембранного потенциала падает в e раз.
2. При коротком стимуле емкостная составляющая будет равна нулю и тогда:
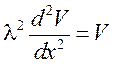 ,
,
где решение: 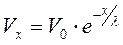 и l – постоянная длины, показывающая через какое расстояние амплитуда мембранного потенциала падает в e раз. С учетом сопротивлений мембраны и аксоплазмы (Rm и Ri):
и l – постоянная длины, показывающая через какое расстояние амплитуда мембранного потенциала падает в e раз. С учетом сопротивлений мембраны и аксоплазмы (Rm и Ri):
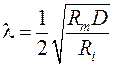 ,
,
где: D – диаметр нервного волокна (D = 2r).
Таким образом, определяющая скорость проведения возбуждения постоянная длины (λ) прямо пропорциональна параметрам нервного волокна.
В итоге, скорость проведения возбуждения:
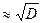 для безмиелинового и
для безмиелинового и 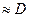 для миелиновых волокон.
для миелиновых волокон.
Из уравнения:
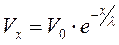
 ,где:
,где:
I0 – начальный ионный ток, RX – входное сопротивление, важное значение для дальнейшей судьбы распространяющегося ПД имеет последняя компонента.
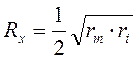
и с учетом удельных значений:
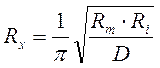 ,
,
где: D – диаметр нервного волокна.
Таким образом, входное сопротивление нервного волокна, от которого зависит скорость распространения нервного импульса, также связана с диаметром этого волокна. Это означает, что с уменьшением диаметра нервного волокна входное сопротивление мембраны значительно возрастает, а при уменьшении – падает. Это явление определяет зависимость скорости проведения возбуждения от геометрических и функциональных неоднородностей нервных волокон. Резкое падение входного сопротивления мембраны будет подобно феномену “перегрузка генератора” в электрической цепи и замедлять проведение возбуждения вплоть до выпадения отдельных нервных импульсов и наоборот – “облегчать” проведение возбуждения при росте входного сопротивления в случае уменьшения диаметра нервного волокна.
Распространение потенциала действия вдоль мембраны нервного волокна будет неодинаково, и из-за этого возникают локальные токи, посредством чего и происходит распространение возбуждения. В каждой точке генерируется новый ПД, однако, проводится не он, а возбуждение в виде локальных токов-пусковых механизмов для новых ПД.
Представим, что в точке VJ мембранный ток (Im) складывается из притекающего и оттекающего токов:
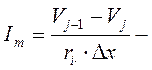
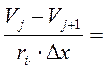
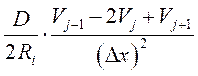 ,
,
где: D – диаметр аксона, Ri- сопротивление аксоплазмы.
Как видно из выражения роль локального тока играет мембранный ток и его характеристики относительно ионных проницаемостей и ПД можно увидеть на Рис.
1. В начале доминирует ток из VJ-1 в VJ, так как первый участок заряжен относительно второго положительно. Этот процесс приводит к деполяризации в точке VJ.
2. В следующей фазе преобладает ток VJ в VJ+1, по выше описанной схеме, так как уже этот участок (VJ) становится положительно заряженным относительно второго.
3. Наконец от возбужденного участка VJ+1 ток течет назад к VJ, вызывая дополнительную деполяризацию мембраны.
На пике первой фазы открываются натриевые каналы, и они забивают мембранный ток, в результате чего он не может внести заметных изменений в Im, и его падение в отрицательную фазу INa замедлено. В результате условием развития ПД будет равенство входящего (INa) и выходящего суммарных токов:
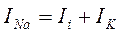
К моменту развития ПД суммарный ионный ток приобретает входящее направление и из уравнения:
,
где в момент развития ПД емкостная составляющая равна нулю и:
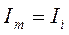 .
.
Эта величина ионного тока получила название ξ-потенциал.
К моменту развития ПД суммарный ионный ток приобретает входящее направление, совпадающее по максимуму с точкой максимальной скорости нарастания ПД. Этот процесс обеспечивает развитие ПД, хотя в точке максимума ПД ионный ток равен нулю. В фазу реполяризации мембраны вновь появляется ионный ток противоположного направления, соответствующий максимальной амплитуде точке максимального снижения ПД.
Дата добавления: 2015-06-22; просмотров: 1832;