Метод електронного парамагнітного резонансу
Явище електронного парамагнітного резонансу(ЕПР) було відкрите радянським фізиком Е. К. Завойським у 1944 році. З того часу цей метод швидко розвинувся і нині став незамінним при вивченні структури вільних органічних та неорганічних радикалів.
Розглянемо систему парамагнітних частинок, парамагнетизм яких зумовлений наявністю неспареного електрона (вільні радикали, іони змінної валентності). Парамагнетизм таких частинок має, як правило, спіновий характер, тобто зумовлений наявністю нескомпенсованого спінового магнітного моменту електрона, який можна записати у вигляді:
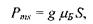 (7.16)
(7.16)
де 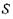 — спінове квантове число (для електрона
— спінове квантове число (для електрона 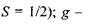 так званий "жи-фактор" (це є безрозмірна величина, яка у разі чисто "спінового магнетизму", тобто для вільного електрона, становить
так званий "жи-фактор" (це є безрозмірна величина, яка у разі чисто "спінового магнетизму", тобто для вільного електрона, становить 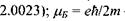 - магнетон Бора.
- магнетон Бора.
При відсутності зовнішнього магнітного поля систему таких парамагнітних частинок можна охарактеризувати деякою середньою енергією 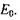 У зовнішньому магнітному полі енергія парамагнітної частинки змінюється і стає рівною
У зовнішньому магнітному полі енергія парамагнітної частинки змінюється і стає рівною
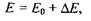 (7.17)
(7.17)
де 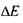 - додаткова енергія, зумовлена взаємодією магнітного моменту частинки з зовнішнім магнітним полем. Ця енергія пропорційна індукції магнітного поля
- додаткова енергія, зумовлена взаємодією магнітного моменту частинки з зовнішнім магнітним полем. Ця енергія пропорційна індукції магнітного поля 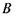 і проекції магнітного моменту
і проекції магнітного моменту 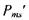 на напрямок поля:
на напрямок поля:
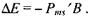 (7.18)
(7.18)
Проекція спінового магнітного моменту на напрямок зовнішнього магнітного поля може набувати значень, що визначаються формулою
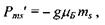 (7.19)
(7.19)
де 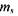 - магнітне спінове число електрона
- магнітне спінове число електрона 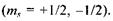 Знак
Знак 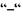 у формулі (7.19) відтворює той факт, що спінові механічний та магнітний моменти спрямовані в протилежні боки.
у формулі (7.19) відтворює той факт, що спінові механічний та магнітний моменти спрямовані в протилежні боки.
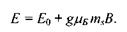 (7.20)
(7.20)
Таким чином, система ізоенергетичних частинок з енергією 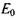 у зовнішньому магнітному полі розпадається на дві підсистеми з енергіями Е1 і Е2 відповідно до двох можливих орієнтацій їх магнітних моментів відносно зовнішнього магнітного поля (рис. 7.5).
у зовнішньому магнітному полі розпадається на дві підсистеми з енергіями Е1 і Е2 відповідно до двох можливих орієнтацій їх магнітних моментів відносно зовнішнього магнітного поля (рис. 7.5).
Між рівнями 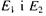 можливі переходи, тобто парамагнітні частинки можуть змінювати орієнтацію свого магнітного моменту. Однак ймовірність спонтанних, самодовільних переходів частинок між рівнями
можливі переходи, тобто парамагнітні частинки можуть змінювати орієнтацію свого магнітного моменту. Однак ймовірність спонтанних, самодовільних переходів частинок між рівнями 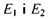 дуже незначна.
дуже незначна.
Такі переходи можна викликати (індукувати) опроміненням системи високочастотним електромагнітним полем (НВЧ-випромінюванням) за умови, що квант НВЧ-випромінювання задовольняє умові 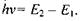
Заселеність енергетичних рівнів 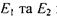 підлягає розподілу Больцмана, згідно з яким при
підлягає розподілу Больцмана, згідно з яким при 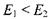 маємо
маємо 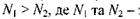 заселеності рівнів
заселеності рівнів 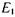 та
та 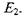 У цьому випадку кількість переходів з поглинанням енергії
У цьому випадку кількість переходів з поглинанням енергії 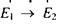 буде пеоеважати кількість переходів з випромінюванням енергії
буде пеоеважати кількість переходів з випромінюванням енергії 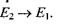 В результаті за умови
В результаті за умови
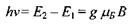 (7.21)
(7.21)
буде спостерігатись резонансне поглинання енергії високочастотного електромагнітного поля (НВЧ-випромінювання).
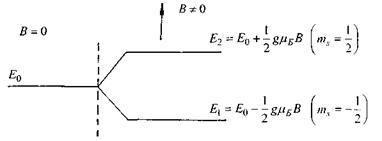
Рис. 7.5. Розщеплення енергетичного рівня парамагнітних частинок.
Для спостереження сигналу поглинання використовуються спеціальні прилади - магнітні радіоспектрометри.
Відповідно до (7.21), умову резонансу можна здійснити двома способами:
а) при постійному магнітному полі 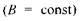 варіюється частота
варіюється частота 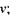
б) при незмінній частоті 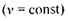 варіюється індукція магнітного поля
варіюється індукція магнітного поля 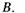
7.3.2. Метод спінових міток (спінових зондів)
Для вивчення структури і функцій біологічних мембран з успіхом застосовується метод спінових міток, або спінових зондів.
Пояснимо суть цього методу. Похідну стійкого вільного радикала, частіше нітроксильного, пов'язують ковалентним зв'язком з будь-якою функціональною групою білка або ліпіду, а потім таку спін-мічену молекулу вбудовують в ліпідний бішар мембран, утворюючи так званий "спіновий зонд". Форма сигналу спінової мітки залежить від обертальної рухливості зонда або тієї його ділянки, до якої вона "пришита" (рис.7.6).

Рис. 7.6. Форми сигналу спінової мітки в залежності від рухливості
зонда.
Використання методу спінових зондів дало можливість встановити цілий ряд цікавих фактів.
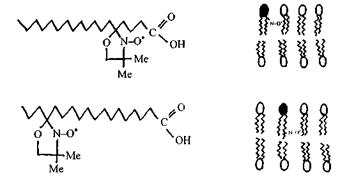
Рис. 7.7. Спінові зонди - спін-мічені аналоги стеаринових кислот.
1. Використовуючи в ролі спінових зондів спін-мічені аналоги стеаринових кислот і вводячи їх в оболонки еритроцитів (рис. 9.28), довели, що відносно жорсткий порядок поширюється на глибину не більш ніж вісім вуглецевих атомів ланцюга від поверхні в глибину мембрани. У напрямку до центра ліпідного бішару рухливість жирно-кислотних ланцюжків зростає. Оцінка обертальної рухливості дала можливість встановити, що мікров'язкість ліпідного бішару в мембрані еритроцита приблизно дорівнює в'язкості рослинної олії.
2. Аналогічним чином, шляхом введення в біологічні мембрани спін-мічених жирних кислот або синтезованих на їх основі фосфоліпідів було виявлено зміну структури мембрани при різних випадках патології, а також під дією ліків та інших біологічно активних сполук. Так, було показано, що анестетики збільшують плинність всередині ліпідного бішару мембрани, а холестерин і ДДТ зменшують її.
3. Методом ЕПР було вперше експериментально доведено існування фосфоліпідного перевороту. Був виявлений повільний фосфоліпідний переворот в мембранних везикулах-ліпосомах, або фосфоліпідних бульбашках (рис. 7.8). Саме таким переворотом можна пояснити механізм пасивного транспорту деяких іонів через біологічні мембрани.

Рис. 7.8. Фосфоліпідний переворот 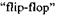 в мембранах.
в мембранах.
Дата добавления: 2015-06-22; просмотров: 1004;