Трансмембранные потоки ионов
Пассивное движение ионов через мембраны (без участия активных транспортных систем) происходит при наличии движущей силы – градиента электрохимического потенциала. Согласно основному уравнению электродиффузионной теории, ионный поток пропорционален движущей силе: 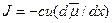 , где c – концентрация иона в данной области, u – подвижность иона в данной среде, а
, где c – концентрация иона в данной области, u – подвижность иона в данной среде, а 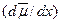 – градиент электрохимического потенциала для данного вида ионов. Трансмембранный градиент электрохимического потенциала для ионов j представляют в виде:
– градиент электрохимического потенциала для данного вида ионов. Трансмембранный градиент электрохимического потенциала для ионов j представляют в виде:
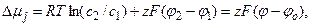 (4.18)
(4.18)
где φ = φ2 – φ1 – мембранный потенциал, а 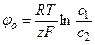 – равновесный потенциал для данного вида ионов. Выражение
– равновесный потенциал для данного вида ионов. Выражение 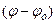 характеризует движущую силу транспорта. Поток ионов J создает электрический ток I:
характеризует движущую силу транспорта. Поток ионов J создает электрический ток I: 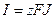 . Коэффициент пропорциональности между величиной тока и действующей силой представляет собой проводимость g:
. Коэффициент пропорциональности между величиной тока и действующей силой представляет собой проводимость g:
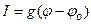 (4.19).
(4.19).
В случае идентичного состава растворов с обеих сторон мембраны, проводимость для иона j равна:
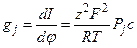 , (4.20)
, (4.20)
где Рj – проницаемость мембраны для данного вида ионов, с – концентрация ионов, а φ – мембранный потенциал. Формула (4.20) описывает зависимость проводимости от концентрации проникающего электролита и проницаемости мембраны. Она не применима для описания изменений проводимости, связанных с переходами каналов между открытым и закрытым состояниями.
Для описания изменений проводимости потенциал-зависимых ионных каналов удобна кинетическая модель с двумя состояниями (закрытым и открытым), переходы между которыми зависят от мембранного потенциала с константами a и b 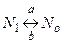 ,
, 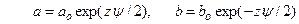 .
.
В данной модели зависимость числа открытых состояний No от потенциала описывается сигмоидной (больцмановской) кривой
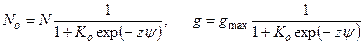 , (4.21)
, (4.21)
где N – общее число каналов, Ko = bo/ao – константа равновесия для переходов между двумя состояниями каналов при отсутствии мембранного потенциала, а ψ=φF/(RT) – мембранный потенциал в единицах RT/F (≈ 25 мВ при 20˚С).
Величина тока в системе потенциал-зависимых каналов определяется произведением общего числа каналов N, вероятности открытого состояния p=No/N, а также тока через одиночный канал i: 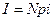 , где i = g(φ – φo) – ток одиночного канала. Из уравнений (4.19, 4.21) видно, что изменения мембранного потенциала оказывают влияние как на движущую силу, так и на проводимость каналов. Поэтому, зависимость тока от напряжения на мембране в системе управляемых потенциалом каналов проявляет существенную нелинейность и может включать участок отрицательного наклона.
, где i = g(φ – φo) – ток одиночного канала. Из уравнений (4.19, 4.21) видно, что изменения мембранного потенциала оказывают влияние как на движущую силу, так и на проводимость каналов. Поэтому, зависимость тока от напряжения на мембране в системе управляемых потенциалом каналов проявляет существенную нелинейность и может включать участок отрицательного наклона.
Участок с отрицательным наклоном характерен для возбудимых мембран. Такая вольт-амперная характеристика (ВАХ) появляется при условии, что область изменений проводимости g(φ), определяемая уравнением (4.21), сдвинута в сторону более отрицательных потенциалов по сравнению с равновесным потенциалом φo для данного вида ионов. Поэтому ВАХ для тока натриевых каналов (φo ~ +50 мВ) включает участок отрицательного сопротивления, а ВАХ для тока калиевых каналов в обычных условиях (φo < –100 мВ) не содержит участка отрицательного наклона.
Пример 4.7. Изобразить зависимость тока через Na канал от мембранного потенциала в интервале от –100 до 100 мВ при условии, что канал с проводимостью 5 пСм открывается при деполяризации мембраны до уровня φ > –50 мВ, а наружная и внутренняя концентрации Nа+ составляют соответственно 145 и 12 мМ. Температура 37°С.
Решение: В области –100 до –50 мВ канал закрыт, ток равен нулю. В области от –50 до 100 мВ зависимость линейна i = g(φ – φo), где φo=(RT/F)·ln(co/ci)= 66,5 мВ. Положительные значения тока при φ > φo соответствуют выходящему направлению (из клетки в среду), а отрицательные токи при φ < φo соответствуют входящему току. При потенциале φ = 0 величинаNa+-тока составит i ≈ 5·10–12 См·(–0,067 В) ≈ -0,33 пА. При потенциале открывания канала φ = –50 мВ, i ≈ 5·10–12·(–0,05–0,067) ≈ -0,585 пА.
Суммарный ток, протекающий через мембраны, включает емкостную составляющую и ионный ток, состоящий из токов отдельных видов ионов: 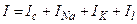 , где Il – ток утечки, для которого характерна линейная зависимость от напряжения на мембране. В физиологических условиях суммарный ток не выводится во внешнюю измерительную цепь и (при разомкнутой внешней цепи) равен нулю. В этих условиях ионный ток равен по абсолютной величине емкостному току.
, где Il – ток утечки, для которого характерна линейная зависимость от напряжения на мембране. В физиологических условиях суммарный ток не выводится во внешнюю измерительную цепь и (при разомкнутой внешней цепи) равен нулю. В этих условиях ионный ток равен по абсолютной величине емкостному току.
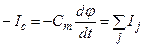 (4.22)
(4.22)
Из (4.22) видно, что перенос ионов через мембрану вызывает изменение потенциала на мембране, представляющей диэлектрический слой (конденсатор) с емкостью Сm. В случае переноса определенного количества зарядов ΔQ (ΔQ = ∫I·dt) изменение напряжения Δφ составит:
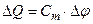 (4.23)
(4.23)
Эта формула позволяет оценить мембранный потенциал, создаваемый при переносе известного количества зарядов, или же оценить минимально необходимый ионный поток сдвигающий напряжение на мембране в период потенциала действия. По данным о количестве перенесенных зарядов можно оценить изменения ионных концентраций.
Пример 4.8. Найти минимальное количество n молей Сa2+, которое необходимо перенести через поверхность мембраны нейрона (в расчете на 1 см2) для генерации потенциала действия с амплитудой 100 мВ. Емкость мембраны Сm = 1 мкФ/см2. Насколько изменится внутриклеточная [Сa2+] при переносе найденного количества Са2+ во время потенциала действия в нейроне диаметром 50 мкм? При расчете допустить, что клетка – сферическая, а внутриклеточный уровень [Са2+] в покое составляет 100 нМ.
Решение: Найдем заряд, который необходимо перенести через мембрану, чтобы сдвинуть потенциал на 100 мВ: 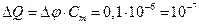 Кл/см2. Пересчитаем количество перенесенных зарядов на соответствующее количество молей вещества (z = 2)
Кл/см2. Пересчитаем количество перенесенных зарядов на соответствующее количество молей вещества (z = 2) 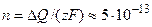 моль/см2. Изменение концентрации внутри клетки при переносе внутрь количества Са2+, равного n, составит
моль/см2. Изменение концентрации внутри клетки при переносе внутрь количества Са2+, равного n, составит 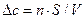 , где
, где 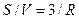 (см пример 4.1):
(см пример 4.1): 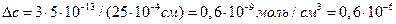 М. Расчет показывает, что поток Са2+ во время потенциала действия вызывает многократное повышение исходного уровня Са2+.
М. Расчет показывает, что поток Са2+ во время потенциала действия вызывает многократное повышение исходного уровня Са2+.
Как видно из (4.22), емкость мембраны определяет скорость изменения мембранного потенциала за счет движения ионов по градиенту электрохимического потенциала. При подключении заряженного конденсатора C к резистору R в аналоговой модели мембраны, напряжение на конденсаторе снижается экспоненциально с постоянной времени τ = RC:
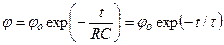 . (4.24)
. (4.24)
Поскольку удельная емкость биологических мембран Сm не претерпевает существенных изменений и составляет по разным оценкам от 0,5 до 1,0 мкФ/см2, анализ кинетики релаксации мембранного потенциала хлоропластов после короткой заряжающей мембрану вспышки света позволяют оценить проводимость тилакоидных мембран в различных условиях (на свету и в темноте, при действии мембрано-активных агентов, при разных концентрациях ионов в среде):
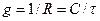 (4.25)
(4.25)
Размерность правой части (4.25) – сименс: [Ф/с] = [Кл/(В с)] = [А/В] = [Ом–1] = [См]. Во многих случаях размерности g и C отнесены к единице площади мембраны [Ф/см2] и [См/см2]. Сочетание (4.20) и (4.25) позволяет иногда оценивать проницаемость мембран по измерениям τ.
Пример 4.9. В приближении постоянного поля оценить удельные значения проводимости и сопротивления тилакоидной мембраны хлоропласта, окруженной растворами с одинаковым содержанием KCl (0,1 М), при условии, что основной вклад в проводимость связан с высокой проницаемостью мембран для К+ (РК = 4·10-8 см/с). Какова постоянная времени мембраны t, если удельная емкость мембраны составляет 1 мкФ/см2?
Решение: Проводимость мембраны оценивают по формуле (4.20), а емкость – по (4.25):
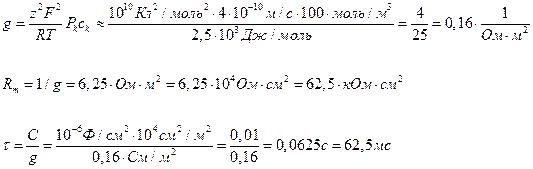
Пример 4.10. При измерениях кинетики спада индуцируемого вспышкой мембранного потенциала Δφ в тилакоидах хлоропластов найдено, что постоянная времени τ экспоненциальной кривой составляет после темновой адаптации хлоропластов 100 мс, а после предварительного освещения – 50 мс. В присутствии ингибитора Н+-АТФазы предварительное освещение хлоропластов не вызывало ускорения спада Δφ. Исходя из допущения, что снижение τ после предварительного освещения обусловлено активацией проводимости сопрягающего фактора CF1-CF0, оценить проводимость одиночного канала Н+-АТФазы и стационарный поток Н+ через одиночный канал при ΔрН ~ 3,3. Плотность расположения Н+-АТФаз в мембране тилакоидов составляет 0,2∙10–13 моль/см2.
Решение: Приращение проводимости, связанное по условию задачи с подключением на свету каналов Н+-АТФазы, составит:
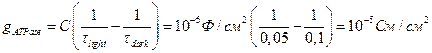
Проводимость одиночного канала Н+-АТФазы находим с учетом плотности расположения АТФаз и числа Авогадро (~6·1023):
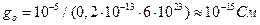 .
.
Для оценки потока протонов выразим протондвижущую силу в единицах напряжения:
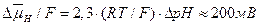 .
.
Ток через одиночный канал составит 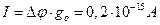 , а количество переносимых H+ составит
, а количество переносимых H+ составит 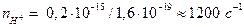 . При стехиометрическом соотношении H+/АТФ = 3, поток H+ соответствует скорости синтеза АТФ ~ 400 c–1 на один комплекс, что соответствует времени оборота фермента ~1/400 = 2,5 мс.
. При стехиометрическом соотношении H+/АТФ = 3, поток H+ соответствует скорости синтеза АТФ ~ 400 c–1 на один комплекс, что соответствует времени оборота фермента ~1/400 = 2,5 мс.
Дата добавления: 2015-06-22; просмотров: 2750;