Передвижение воды по растению.
Вода, поступившая в клетки корня под влиянием разности водных потенциалов, которые возникают благодаря транспирации и корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе может перемещаться в радиальном направлении тремя путями: апопластическим, симпластическим, трансмембранным. Еще в 1932 г. немецкий физиолог Э. Мюнх высказал мнение о существовании в корневой системе двух относительно независимых друг от друга объемов, по которым передвигается вода,— апопласта и симпласта. При транспорте по апопласту вода передвигается по клеточным стенкам, не проходя через мембраны. При симпластном транспорте вода проникает в клетку через полупроницаемую мембрану и далее перемещается по протопластам клеток, которые соединены между собой многочисленными плазмодесмами. При трансмембранном транспорте вода перетекает через клетки и при этом проходит, по крайней мере, две плазматические мембраны. Уже обсуждалось, что в последнее время много внимания уделяется аквапоринам — мембранным белкам, образующим в мембранах специализированные водные каналы и определяющим проницаемость для воды.
Эксперименты показали, что передвижение воды по коре корня идет главным образом по апопласту, где она встречает меньшее сопротивление, и лишь частично по симпласту (С. Френч). Апопластный путь прерывается в эндодерме в связи с наличием поясков Каспари. Вместе с тем в апикальной части суберинизация отсутствует, поэтому вода легко проникает через эндодерму. Кроме того, в суберинизированных частях корня вода может проходить через пропускные клетки.
Сказанное показывает, что для транспорта в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану клеток эндодермы. Таким образом, мы имеем дело как бы с осмометром, у которого полупроницаемая мембрана расположена в клетках эндодермы. Вода устремляется через эту мембрану в сторону меньшего (более отрицательного) водного потенциала. Далее вода поступает в сосуды ксилемы. По вопросу о причинах, вызывающих секрецию воды в сосуды ксилемы, имеются различные суждения. Согласно гипотезе Крафтса, это следствие выброса солей в сосуды ксилемы, в результате чего там создается повышенная их концентрация, и водный потенциал становится более отрицательным. Предполагается, что в результате активного поступления соли накапливаются в клетках корня. Однако интенсивность дыхания в клетках, окружающих сосуды ксилемы (перицикл), очень низкая, и они не удерживают соли, которые благодаря этому десорбируются в сосуды.
Транспорт воды в корне зависит от интенсивности процесса дыхания. При помещении растений в условия, тормозящие дыхание корней (низкая температура, анаэробиоз или наличие дыхательных ядов), они транспортируют меньше воды. Предполагают, что это может быть связано с инактивированием аквапоринов. Торможение транспорта воды в корнях в аэробных условиях, возможно, объясняет факт завядания растений в переувлажненной почве.
Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов и трахеид.
Опыты с кольцеванием показали, что восходящий ток воды по растению движется в основном по ксилеме. В проводящих элементах ксилемы вода встречает незначительное сопротивление, что, естественно, облегчает передвижение воды на большие расстояния. Правда, некоторое количество воды передвигается и вне сосудистой системы. Однако по сравнению с ксилемой сопротивление движению воды других тканей значительно больше. Это приводит к тому, что вне ксилемы движется всего от 1 до 10% общего потока воды.
Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводя-щие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры — отличительной чертой растений, устойчивых к засухе.
Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. В связи с этим, как только в силу процесса транспирации возникает ненасыщенность водой клеточных стенок паренхимных клеток, она сейчас же передается внутрь клетки, водный потенциал которой падает. Вода передвигается от клетки к клетке благодаря градиенту водного потенциала. По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам, где сопротивление значительно меньше.
По сосудам вода движется благодаря создающемуся в силу транспирации градиенту водного потенциала, градиенту свободной энергии (от системы с большей свободой энергии к системе с меньшей). Можно привести примерное распределение водных потенциалов, которое и вызывает передвижение воды: водный потенциал почвы (-0,5 бар), корня (-2 бар), стебля (-5 бар), листьев (-15 бар), воздуха при относительной влажности 50% (-1000 бар).
Однако получены экспериментальные данные, которые не позволяют рассматривать силу транспирации как единственную, обуславливающую восходящий ток воды по растению. Так, показано, что восходящий ток воды может осуществляться и при отсутствии транспирации. К этому же выводу приводят опыты, показывающие ритмическое секретирование устьичными клетками жидкой воды, а также зависимость передвижения воды от эндогенной энергии, поставляемых процессом дыхания. Это позволяет считать, что движущая сила транспорта воды в растении является суммой двух весьма различных по своей природе составляющих, условно названных метаболической и осмотической. Об этом уже упоминалось при рассмотрении вопроса о корневом давлении. Осмотическая составляющая представлена в корнях сугубо осмотическими явлениями, в стебле и листьях — гидростатической тягой, создаваемой градиентом водного потенциала в системе почва — растение — атмосфера. Температурный коэффициент Q10 этой составляющей близок к единице. Именно такая величина Q10 характерна для простых физических процессов. В то же время Q10 процесса транспорта воды значительно выше. Столь высокий температурный коэффициент свойственен сложным процессам цепного характера. Очевидно, транспорт воды за счет метаболической составляющей относится к разряду именно таких процессов. Согласно имеющимся экспериментальным данным, в формировании метаболической составляющей непосредственное участие принимают контрактильные системы паренхимных клеток (т. е. белки цитоскелета). Они могут играть роль сфинктеров, регулирующих просветы водных каналов (открывание—закрывание) в плазмодесмах. Благодаря их работе индуцируются ритмические микроколебания внутриклеточного (внутрисимпластного) гидростатического давления, которые внешне выражаются в короткопериодных (1—3 мин) автоколебаниях поступления воды в клетку (симпласт, ткань, орган) и водоотдачи (т. е. микропульсациях). Принципиально важное значение имеет факт противофазное™ автоколебаний водопоглощения и водоотдачи. Это свидетельствует о том, что процесс транспорта воды состоит из двух последовательных, ритмично чередующихся, относительно самостоятельных, хотя и тесно взаимосвязанных фаз: фазы сокращения, во время которой вода выделяется (выталкивается, секретируется) и последующей фазы расслабления, во время которой вода поглощается. Выделение воды вызывает падение тургора и водного потенциала в целом, создавая предпосылку для поглощения следующей порции воды, вновь приводящего к возрастанию водного потенциала вплоть до того, что он из отрицательного становится положительным. После этого происходит новое сокращение. Именно фаза сокращения происходит с участием контрактильных систем и требует затраты энергии. Таким образом, вода поглощается и выделяется по градиенту водного потенциала, а не против него, т. е. согласно данной схеме, транспорт воды в термодинамическом понимании является пассивным. Возникающие за счет ритмической деятельности внутриклеточного сократительного аппарата микроколебания гидростатического давления паренхим-ных клеток являются механизмом, создающим локальные градиенты водного потенциала на пути водного тока и тем самым регулирующим скорость этого тока. Именно благодаря этому формируется метаболическая составляющая движущей силы транспорта воды в растении, играющая решающую роль в общей системе эндогенной регуляции. Под влиянием ингибиторов контрактильных систем или окислительного фосфорилирования {т. е. при нарушении энергоснабжения) противофазность исчезает, автоколебания затухают и транспорт воды тормозится (В.Н. Жолкевич).
Необходимо также отметить роль сил сцепления для обеспечения поднятия воды в растении на высоту более 100 м. Теория сцепления была выдвинута русским ученым Е.Ф. Вотчалом и английским физиологом Е. Диксоном. Для лучшего понимания рассмотрим следующий опыт. В чашку с ртутью помещают заполненную водой трубку, которая заканчивается воронкой из пористого фарфора. Вся система лишена пузырьков воздуха. По мере испарения воды ртуть поднимается по трубке. При этом высота подъема ртути превышает 760 мм. Это объясняется наличием сил сцепления между молекулами воды и ртути, которые в полной мере проявляются при отсутствии воздуха. Сходное положение, только еще более ярко выраженное, имеется в сосудах у растений.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту, значительно большую Юм. Расчеты показали, что благодаря наличию сродства между молекулами воды силы сцепления достигают величины —30 бар. Это такая сила, которая позволяет поднять воду на высоту, равную 120 м, без разрыва водных нитей, что примерно составляет максимальную высоту деревьев. Силы сцепления существуют и между водой и стенками сосудов — адгезия. Стенки проводящих элементов ксилемы эластичны. В силу этих двух обстоятельств даже при недостатке воды связь между молекулами воды и стенками сосудов не нарушается. Это подтверждается исследованиями по изменению толщины стебля травянистых растений. Определения показали, что в полуденные часы толщина стебля растений уменьшается. Если перерезать стебель, то сосуды сразу расширяются и воздух поступает в них. Из этого опыта видно, что при сильном испарении сосуды сужаются и это приводит к появлению отрицательного давления.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую кашпо испаряемой воды. Таким образом, при нормальном водоснабжении создается непрерывность воды в системе почва — растение — атмосфера. В том случае, если в отдельные членики сосудов попадает воздух (эмболия), они, по-видимому, выключаются из общего тока проведения воды. Таков путь воды по растению и его основные движущие силы.
Современные методы исследования позволяют определить скорость передвижения воды по растению. Скорость передвижения воды определяется разностью водных потенциалов в начале и конце пути, а также сопротивлением, которое она встречает. Было предложено распространить на этот процесс закон Ома, тогда
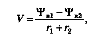
где V— скорость движения воды, Ψв1 и Ψв2 — водные потенциалы, r1 и r2— сопротивления. Согласно полученным данным, скорость движения воды в течение суток изменяется и в дневные часы она значительно больше. При этом разные виды растений отличаются по скорости передвижения воды. Если скорость передвижения у хвойных пород обычно 0,5—1,2 м/ч, то у лиственных она значительно выше. У дуба, например, скорость передвижения составляет 27 — 40 м/ч. Скорость передвижения воды мало зависит от напряженности обмена веществ. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды скорее могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Дата добавления: 2015-06-05; просмотров: 2484;