Расходование воды растением – транспирация.
В основе расходования воды растительным организмом лежит процесс испарения — переход воды из жидкого в парообразное состояние, происходящий при соприкосновении органов растения с ненасыщенной водой атмосферой. Однако этот процесс осложнен физиологическими и анатомическими особенностями растения, и его называют транспирацией.
Количество воды, испаряемой растением, во много раз превосходит объем содержащейся в нем воды. Экономный расход воды составляет одну из важнейших проблем сельскохозяйственной практики. К.А. Тимирязев назвал транспирацию в том объеме, в каком она идет, «необходимым физиологическим злом». Действительно, в обычно протекающих размерах транспирация не является необходимой. Так, если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти со значительно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Вместе с тем транспирация в определенном объеме полезна растительному организму:
1. Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может примерно на 7°С быть ниже температуры листа завядающего, нетранспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза 20—25°С). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2. Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
3. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом чем интенсивнее транспирация, тем быстрее идет этот процесс.
Как уже говорилось, механизм поступления ионов и воды в клетку различен. Однако некоторое количество питательных веществ может поступать пассивно, и этот процесс может ускоряться с увеличением транспирации.
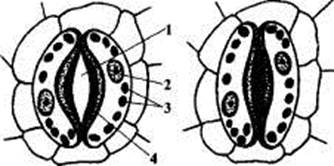
Строение листа. Устьица.Основным транспирирующим органом является лист. Средняя толщина листа составляет 100—200 мкм. Паренхимные клетки листа расположены рыхло, между ними имеется система межклетников, которые занимают от 15 до 25% объема листа. Эпидермис — покровная ткань листа, состоит из компактно расположенных клеток, наружные стенки которых утолщены. Кроме того, листья большинства растений покрыты кутикулой, в состав которой входят оксимонокарбоновые кислоты, содержащие по 16—28 атомов углерода и по 2—3 гидроксильные группы. Эти кислоты соединены друг с другом в цепочки с помощью эфирных связей. Кутикула варьирует как по составу, так и по толщине. Более развитой кутикулой характеризуются листья светолюбивых растений по сравнению с теневыносливыми и засухоустойчивых по сравнению с влаголюбивыми. Кутикула вместе с клетками эпидермиса образует как бы барьер на пути испарения паров воды. Удаление кутикулы во много раз повышает интенсивность испарения. Все эти особенности выработались в процессе эволюции как приспособление к сокращению испарения. Для соприкосновения листа с атмосферой имеются поры — устьица. Устьице — это отверстие (щель), ограниченная двумя замыкающими клетками. Устьица встречаются у всех наземных органов растения, но больше всего у листьев. Каждая замыкающая клетка устьица в отличие от клеток эпидермиса имеет хлоропласты. В них происходит фотосинтез, хотя с меньшей интенсивностью, чем в клетках мезофилла. Устьица — одно из оригинальных приспособлений, обладающих способностью открываться и закрываться в зависимости от насыщенности замыкающих клеток водой. Обычно устьичные отверстия ограничены двумя замыкающими клетками, стенки которых неравномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу клеточные стенки более толстые, а внешние — более тонкие. Протопласты замыкающих клеток связаны в единое целое перфорациями в основании граничащих общих стенок. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на клеточные стенки, и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель (рис. 1).
|
Рисунок 1 - Структура устьиц у двудольных растений:
А — открытое устьице;
Б — закрытое устьице;
1 — устьичная щель;
2 — ядро;
3 — хлоропласты;
4 — толстая клеточная оболочка.
В последнее время доказано, что для движения устьиц большое значение имеет также расположение микрофибрилл целлюлозы. Если обычно в клетках листьев целлюлозные фибриллы ориентированы в длину и в этом направлении утолщены, то в замыкающих клетках устьиц микрофибриллы организованы радиально, что усиливает устойчивость к процессу растяжения.
У злаков строение замыкающих клеток несколько иное. Они представлены двумя удлиненными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель (рис. 2).
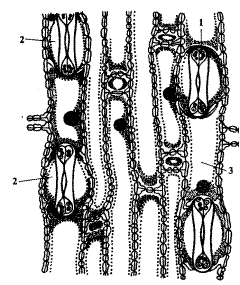
Рисунок 2 - Структура устьиц у однодольных растений
1 — замыкающие клетки устьиц; 2 — побочные клетки; 3 — клетки эпидермиса
Число устьичных отверстий колеблется в зависимости от вида растений от 10 до 600 на 1 мм2 листа. У многих растений (75% видов), в том числе для большинства древесных, устьица расположены на нижней стороне листа. Диаметр устьичных щелей составляет всего 3—12 мкм. Устьица соединяют внутренние пространства листа с внешней средой. Вода поступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны три пути испарения: через устьица — устьичная, кутикулу — кутикулярная и через чечевички — лентикулярная транспирация. Впервые разграничение на кутикулярную и устьичную транспирацию было введено в 1877 г.
Кутикулярная транспирация. Втом, что действительно испарение идет не только через устьица, но и через кутикулу, легко убедиться. Так если взять листья, у которых устьица расположены только с нижней стороны (например, листья яблони), и замазать эту сторону вазелином, то испарение воды будет продолжаться, хотя и в значительно меньших размерах. Следовательно, определенное количество воды испаряется через кутикулу. Интенсивность этого процесса прежде всего определяется толщиной слоя кутикулы. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом. Однако у растений, листья которых характеризуются слабым развитием кутикулы, доля этого вида транспирации может повышаться до 30%. Имеет значение также возраст листа. Молодые листья, как правило, имеют слабо развитую кутикулу и, следовательно, более интенсивную кутикулярную транспирацию. У старых листьев доля кутикулярной транспирации снова возрастает, так как, хотя кутикула и сохраняет достаточную толщину, в ней появляются трещины, через которые легко проходят пары воды.
Трещины в кутикуле могут появляться и после временного завядания листьев, благодаря чему транспирация усиливается. Кутикулярная транспирация зависит от оводненности листа. При насыщении кутикулы водой испарение идет интенсивнее, а при подсыхании кутикулы — снижается. Имеются данные, что кутикулярная транспирация меньше зависит от условий внешней среды по сравнению с устьичной (Н.И. Антипов).
Устьичная транспирация.Основная часть воды испаряется через устьица. Процесс транспирации можно разделить на ряд этапов.
Первый этап — это переход воды из клеточных оболочек, где она находится в капельножидком состоянии, в межклетники (парообразное состояние). Это собственно процесс испарения, отрыв молекул воды с поверхности клеточных стенок. Важно подчеркнуть, что уже на этом этапе растение обладает способностью регулировать процесс транспирации (внеустьичная регулировка). Так, если в растении недостаток воды, то в сосудах корня и стебля создается сильное натяжение, которое делает их водный потенциал более отрицательным, что оказывает сопротивление передвижению воды в клетку и уменьшает интенсивность испарения. Надо учитывать также, что между всеми частями клетки существует водное равновесие. Чем меньше воды в клетке, тем выше становится концентрация клеточного сока. А это, в свою очередь, уменьшает содержание свободной воды в протопласте и клеточной оболочке. Соотношение свободной воды к связанной падает, водоудерживающая сила растет, интенсивность испарения уменьшается. Кроме того, снижение оводненности клеточных стенок приводит к изменению формы менисков в капиллярах на вогнутую. Это увеличивает поверхностное натяжение, затрудняет переход воды в парообразное состояние и снижает количество водяного пара в межклетниках.
Второй этап — это выход паров воды из межклетников или через кутикулу, или, главным образом, через устьичные щели. Поверхность всех клеточных стенок, соприкасающихся с межклетными пространствами, превышает поверхность листа примерно в 10—30 раз. Все же если устьица закрыты, то все это пространство быстро насыщается парами воды и переход воды из жидкого в парообразное состояние прекращается. Иная картина наблюдается при открытых устьицах. Как только часть паров воды выйдет из межклетников через устьичные щели, так сейчас же этот недостаток восполняется за счет испарения воды с поверхности клеток. Поскольку устьичная транспирация составляет 80—90% от всего испарения листа, то степень открытости устьиц является основным механизмом, регулирующим интенсивность транспирации. При открытых устьицах общая поверхность устьичных щелей составляет всего 1—2% от площади листа. Казалось бы, это должно очень сильно уменьшать испарение по сравнению с испарением со свободной водной поверхности той же площади, что и лист. Однако это не так. Сравнение испарения листа с испарением со свободной водной поверхности той же площади показало, что оно идет не в 100 раз, как это следовало бы, исходя из размеров открытой площади (1%), а всего в 2 раза медленнее. Объяснение этому явлению было дано в исследованиях английских ученых Г. Броуна и Ф. Эскомба, которые установили, что испарение из ряда мелких отверстий идет быстрее, чем из одного крупного той же площади. Это связано с явлением краевой диффузии. При диффузии из отверстий, отстоящих друг от друга на некотором расстоянии, молекулы воды, расположенные по краям, рассеиваются быстрее. Естественно, что таких краевых молекул значительно, больше в ряде мелких отверстий по сравнению с одним крупным. Для малых отверстий интенсивность испарения пропорциональна их диаметру, а не площади (закон Й. Стефана).
Указанная закономерность проявляется в том случае, если мелкие поры расположены достаточно далеко друг от друга. Структура листа удовлетворяет указанным требованиям. Поры (устьица) имеют малый диаметр и достаточно удалены друг от друга. При открытых устьицах выход паров воды идет достаточно интенсивно, закрытие устьиц резко тормозит испарение. Именно на этом этапе вступает в действие устьичная регулировка транспирации. При недостатке воды в листе устьица автоматически закрываются. Полное закрывание устьиц сокращает транспирацию на 90%. Вместе с тем уменьшение размеров устьиц всегда приводит к соответственному сокращению транспирационного процесса. Определения показали, что устьица должны закрыться больше чем на 1/2 чтобы это сказалось на уменьшении интенсивности транспирации.
Третий этап транспирации — это диффузия паров воды от поверхности листа в более далекие слои атмосферы. Этот этап регулируется лишь условиями внешней среды.
Дата добавления: 2015-06-05; просмотров: 2805;