ВОЗДЕЙСТВИЯ 2 страница
Если предположить возможность сужения капиллярного русла в отделах мозгового вещества с пониженной функциональной деятельностью, сразу же возникает вопрос, насколько велика степень и продолжительность сужения капилляров.
Отсутствие фактических данных заставляет искать ответы на эти вопросы в будущих исследованиях, крайне необходимых для понимания циркуляции крови в мозгу.
Подводя итог всему сказанному, нужно отметить, что циркуляция крови в мозгу значительно отличается от циркуляции ее в ряде органов и прежде всего в мышцах.
В противоположность тому, что наблюдается в мышцах, в мозгу в нормальном состоянии все капилляры открыты. Лишь очень редко встречаются суженные, а возможно, и закрытые капилляры. Усиление функции того или иного отдела мозга сопровождается расширением капилляров в данной области. Возможно, что в участках мозга, функциональная деятельность клеток которых понижена, имеет место сужение капиллярного русла или полное закрытие отдельных капилляров и выключение их из капиллярной сети. Окончательное представление о циркуляции крови в мозгу нормального животного при раздражении различных рецепторов будет получено только после дальнейших экспериментальных исследований.
Мы уже видели, что регистрация состояния сосудистой сети в мягкой мозговой оболочке может быть произведена при помощи непосредственного наблюдения сосудов на поверхности мозга. Метод «окна», герметически вставленного в череп, позволяет проследить реакции сосудов в условиях, в наибольшей степени приближающихся к нормальным, имеющим место в закрытом черепе. В настоящее время этот метод, предложенный Равина еще в 1811 г., усовершенствован применением капилляроскопа, позволяющим добавить к субъективному наблюдению объективное намерение сосудов мягкой мозговой оболочки.
Однако достоинства этого метода, заключающиеся в наглядности наблюдения и легкости документации с помощью фотографии, снижаются тем, что он дает возможность проследить лишь колебания просвета сосудов, проходимых для тока крови. Полностью закрытые сосуды этим методом не отмечаются. Ограниченность применения метода наблюдения сосудов мягкой мозговой оболочки через окно заключается еще и в том, что при пользовании им необходимо учитывать, насколько состояние сосудистой сети в мягкой мозговой оболочке отражает состояние сосудов расположенного под ней мозгового вещества.
Установить взаимные отношения реакций тех и других сосудов необходимо потому, что общий характер сосудистой сети мягкой мозговой оболочки и внутримозгового вещества различен. Например, у человека основную массу сосудов в артериальной сети мягкой мозговой оболочки составляют артерии диаметром от 3 000 до 40 . Лишь сравнительно в
влияний в процессах эмболической закупорки сосудов мозга. Согласно мнению этих авторов, постоянные рефлекторные влияния, исходящие из участка сосуда, закрытого тромбом или эмболом, вызывают значительно большее расстройство кровообращения, чем механическое прекращение тока крови в закрытом сосуде. Иначе говоря, длительное повторение непродолжительного самого по себе спазма ведет к такому же тяжелому нарушению жизнедеятельности нервной ткани, как и полное прекращение кровообращения в каком-либо участке ее.
При изучении характера реакций артерий мягкой мозговой оболочки в ответ на воздействие на них различных факторов особое внимание уделялось влиянию со стороны вегетативного отдела нервной системы.
Теоретический интерес, проявляемый к этой стороне проблемы мозговой циркуляции, объясняется тем значением, какое имеет этот отдел нервной системы в клинике. Известны и широко распространены вмешательства на вегетативной нервной системе при ряде болезней и патологических состояний: гипертонии, эпилепсии, язве желудка, облитериру-ющем эндартериите и др.
И с этой точки зрения понятен тот интерес, который проявляется до наших дней к вопросам, трактующим о взаимодействии нервной и сосудистой системы мозга и об участии нервной системы в регуляции мозгового кровообращения.
Как следует из данных, собранных в главе, освещающей вопросы иннервации сосудов мозга, гистологически может считаться доказанным существование большого количества нервов, сопровождающих сосуды мягкой мозговой оболочки. Наличие нервов, сопровождающих сосуды, говорит, по-видимому, о зависимости просвета артерий от влияний, исходящих от нервной системы. Однако результаты работ, проделанных с помощью физиологических методов исследования, указывают на то, что такое заключение может быть сделано лишь при учете целого ряда ограничений.
Начнем с наблюдений, касающихся реакций артерий мягкой мозговой оболочки при воздействиях на симпатическую нервную систему.
Опыты, проделанные Форбсом, Коббом и другими (Forbes, Cobb, 1928—1939) на большом количестве животных, показали, что раздражение симпатического нерва на шее имеет своим следствием сужение артерий мягкой мозговой оболочки. Эффект раздражения симпатического нерва может быть обнаружен на артериях, диаметр которых превышает 50 м, и с особой отчетливостью выступает у артерий калибром в 110—340 м. Реакция обнаруживается на стороне раздражения и не зави-сит от изменений общего кровяного давления. Последнее обстоятельство послужило поводом к предположению о воздействии симпатической нервной системы непосредственно на стенку мозгового сосуда в смысле изменения ее тонического состояния.
Разбираемые опыты интересны с двух точек зрения.
Во-первых, характерно, что сокращение под влиянием сосудосуживающего импульса симпатической природы было получено лишь у артерий определенного размера. Сокращались артерии того диаметра, для которого гистологически было установлено наличие в адвентиции большого количества нервных волокон. С этой позиции результаты опытов Форбса кажутся весьма доказательными. Вторым интересным фактом, полученным в разбираемой работе, является незначительная величина, на которую изменяют свой просвет артерии мягкой мозговой оболочки при раздражении симпатического нерва. Действительно, если в тех же условиях опыта артерии кожи сокращаются на 50% от исходного
диаметра, артерии мягкой мозговой оболочки при раздражении симпатического нерва уменьшают свой просвет только на 8—10% (рис, 132).
Последнее обстоятельство говорит о незначительной степени влияния со стороны симпатической нервной системы на артерии мягкой мозговой оболочки, находящиеся под ее контролем.
Мы неоднократно имели возможность убедиться в аналогичном действии симпатического нерва на крупные артерии мягкой мозговой оболочки. Используя метод вставления окна в череп при соблюдении полной герметичности, мы проследили ответные реакции артерий мягкой мозговой оболочки при раздражении неперерезанного симпатического нерва на шее животного в течение продолжительного времени. При
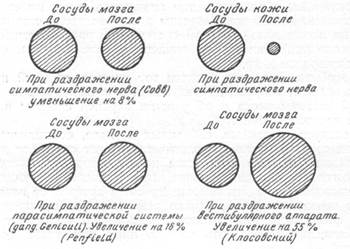
Рис. 132. Изменение диаметра артерий мягкой мозговой оболочки при раздражении симпатической, парасимпатической системы и вестибулярного аппарата (слева — до раздражения; справа — после раздражения).
одновременной записи кровяного давления можно было видеть, что раздражение симпатического нерва не сопровождается изменением величины общего кровяного давления (рис. 133).
На рис. 134, а, б, в, г представлены результаты наблюдений над артериями мягкой мозговой оболочки кошки одновременно с регистрацией общего кровяного давления. Микрофотография (рис, 134, а) была снята в условиях спокойного сна кошки под наркозом. Видна артерия, диаметр которой равняется 140 м, и две расположенные по обе стороны от нее вены. Через 30 секунд после начала раздражения симпатического нерва силой в 13 см индукционной катушки отмечается сужение артерии, просвет которой становится равным 120 м, и в то же время вены сохраняют диаметр, который был до опыта (рис. 134, б). Уменьшенный диаметр артерии сохранялся в данном опыте в продолжение 4—5 минут, но при последующем раздражении размер артерии постепенно возвратился к исходному, а затем превысил его. Так, через 15 минут после начала раздражения симпатического нерва размер артерии, располагавшейся в поле зрения, стал равным 150 м. Вены в продолжение всего указанного времени своих размеров не изменяли (рис. 134, в).
При повторных раздражениях симпатического нерва в одном и том же опыте, произведенных через значительный интервал времени после
первого раздражения, артерии обычно не изменяют своего размера или даже увеличивают его, иногда на значительную величину. Длительное раздражение в этих условиях сопровождается постепенным уменьшением диаметра артерии и возвращением ее величины к исходной, несмотря на существующее раздражение.
Проделанные нами опыты, один из которых здесь приведен, подтверждают данные других исследователей в отношении реакций крупных артерий мягкой мозговой оболочки при раздражении симпатического нерва. Кратковременное раздражение сопровождается уменьшением артерий на величину от 8 до 15% от исходной.
Но длительное раздражение имеет своим следствием или совершенное отсутствие изменения калибра артерии, или искаженную реакцию в виде расширения.
Незначительный предел колебания просвета артерии при раздражении симпатического нерва указывает на слабое влияние со стороны

Рис. 133. Общее кровяное давление у кошки до и после раздражения симпатического
нерва на шее.
а — кровяное давление, записанное до опыта; б — кровяное давление во время раздражения симпатического нерва. Запись сделана ртутным манометром.
симпатического отдела нервной системы на артерии мягкой мозговой оболочки. Венозная сеть мягкой мозговой оболочки, возможно, находится под контролем указанного отдела нервной системы еще в меньшей степени, чем артериальная сеть, и изменения ее не могут быть отмечены имеющимися в нашем распоряжении методами.
В то время как раздражение симпатического нерва влечет за собой сокращение артерий мягкой мозговой оболочки, перерезка симпатического нерва или удаление верхнего шейного узла не ведет, как это можно было ожидать, к расширению артерий [Форбс и Кобб, 1938; Флори, 1925; Ризе (Riser), 1936]. Отсутствие расширения артерий мягкой мозговой оболочки после перерезки симпатических нервов указывает на то, что тоническое состояние сосудистой стенки зависит не только от влияний со стороны симпатического отдела вегетативной нервной системы. Нужно предположить, что влияние со стороны этого отдела вегетативной нервной системы является лишь одним из факторов, совокупность которых определяет тонус стенки сосудов мягкой мозговой оболочки.
При исследовании влияния парасимпатического отдела вегетативной нервной системы на артерии мягкой мозговой оболочки внимание экспериментаторов привлекли блуждающий, лицевой и вестибулярный нервы.
Данные о проведении сосудорасширяющих импульсов по блуждающему нерву были получены в опытах Форбса (1928), который наблюдал расширение артерий мягкой мозговой оболочки при раздражении блуждающего нерва на шее (рис. 135). Опыты Форбса в последующем были повторены Пенфильдом (1932), а также Коббом и Финезингером (1932). Однотипность реакций расширения артерий мягкой мозговой оболочки при раздражении блуждающего нерва, его ветвей или gangl. nodosum в большом количестве опытов на различных животных давала как будто возможность сделать заключение о прохождении сосудорасширяющего импульса именно по этому нерву.
Характерно, что расширение артерий мягкой мозговой оболочки, наблюдаемое на одном -полушарии головного мозга, происходило независимо от того, раздражался блуждающий нерв той же или противоположной стороны. Это явление должно было бы указать авторам на возможность расширения артерий вследствие падения общего кровяного давления. К тому же в большинстве опытов раздражение блуждающих нервов действительно сопровождалось падением общего кровяного давления и только лишь в отдельных экспериментах давление оставалось неизменным. Однако, несмотря на это, Кобб и Финезингер выдвинули ошибочное положение о независимости расширения артерий от падения кровяного давления, т. е. о непосредственном влиянии парасимпатической части вегетативной нервной системы на сосуды мягкой мозговой оболочки.
В экспериментах этих исследователей должна быть отмечена еще одна особенность, заключающаяся в том, что расширение артерий при раздражении блуждающего нерва имело место лишь при сохранении целостности лицевого нерва. Перерезка лицевого нерва не вела к изменению просвета артерий, а стимуляция неизменно сопровождалась расширением их. Исходя из этого факта, Кобб предположил, что сосудорасширяющий импульс для артерий мягкой мозговой оболочки идет по стволу блуждающего нерва на шее. Далее импульс переходит в продолговатом мозгу в область расположения ядер лицевого нерва и по VII паре черепномозговых нервов следует к коленчатому ганглию. От коленчатого ганглия проводящим путем для сосудорасширяющего импульса является уже большой поверхностный каменистый нерв.
Однако указанное положение Кобба не получило подтверждения в работе Форбса и его сотрудников (Forbes, Nason, Wortman), опубликованной в 1937 г. В экспериментах этого исследователя раздражение блуждающего нерва постоянно сопровождалось расширением артерий мягкой мозговой оболочки независимо от сохранности или перерезки лицевого нерва. Тот же самый эффект расширения артерий имел место и при раздражении ветвей блуждающего нерва: нерва каротидного синуса или депрессорного нерва.
В опытах Форбса и сотрудников (1937) раздражение блуждающего нерва или его ветвей во всех случаях сопровождалось также падением общего кровяного давления. Было найдено, что необходимым и обязательным условием для расширения артерий является именно падение общего кровяного давления до низкого уровня (до 60 мм и ниже). Если же во время раздражения блуждающего нерва предотвратить падение давления, то просвет артерий мягкой мозговой оболочки остается неизменным. Кроме того, та же самая реакция со стороны артерий может быть получена и при падении кровяного давления, не связанном с раздражением блуждающего нерва. Так, например, остановка сердечной деятельности, падение давления в мозгу при зажатии обеих сонных арте-
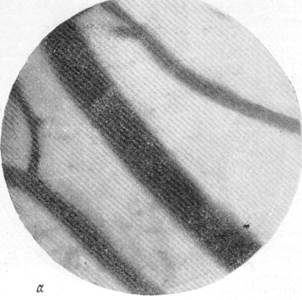
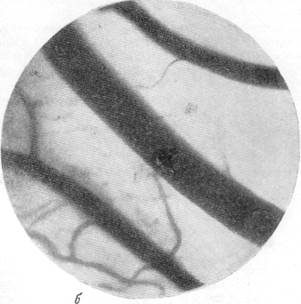
Рис. 134. Реакции артерий и вен мягкой мозговой оболочки при раздражении симпатического нерва.
а — спокойный сон в состоянии наркоза; б — через 30 секунд после начала раздражения
симпатического нерва.
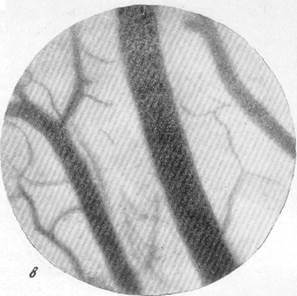
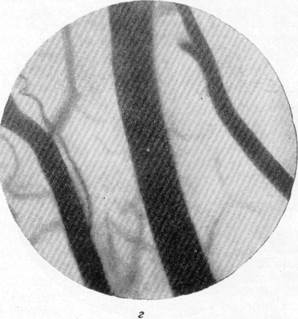
Рис. 134. Реакции артерий и вен мягкой мозговой оболочки при раздражении симпатического нерва.
в — через 15 минут после начала раздражения симпатического нерва; г — через 5 минут после прекращения раздражения. Фото через "окно" в черепе с помощью капилляро-
скопа. Увеличение 60.
рий и т. д. влекут за собой расширение артерий мягкой мозговой оболочки.

|
Приведенные факты указывают лишь на косвенное отношение блуждающего нерва или его ветвей к реакции расширения артерий. Они заставляют предполагать также, что раздражение указанных нервов ведет к падению общего кровяного давления вследствие расширения сосудов в спланхнической области. Результатом резкого падения давления является замедление скорости тока крови в сосудах мозга, расслабление сосудистой стенки и расширение сосудов мягкой мозговой оболочки.
Таким образом, первоначальное предположение о проведении сосудорасширяющего импульса для артерий мозга по блуждающему нерву не получило подтверждения.
На настоящем этапе наших знаний по данному вопросу часть пути сосудорасширяющего импульса с периферии до продолговатого мозга не может считаться установленной и требует дальнейшей экспериментальной разработки.
| Рис. 135. Раздражение блуждающего нерва при перерезке лицевого нерва соответствующей стороны (по Форбсу). В верхней части таблицы приведены изменения диаметра артерий мягкой мозговой оболочки: в нижней части — соответствующие им величины падения общего кровяного давления при раздражении блуждающего нерва. Точкой отмечен нижний уровень давления. |
Как уже указывалось, в работах Пенфильда, Хоробского и Пенфильда, а затем Кобба и Финезингера было установлено, что сосудорасширяющий импульс для артерий мягкой мозговой оболочки идет от продолговатого мозга к узлу колена лицевого нерва (gangl. geniculi n. facialis). Раздражение любой точки на пути этого импульса имеет своим следствием расширение артерий мягкой мозговой оболочки. Но в отличие от блуждающего нерва раздражение лицевого нерва вблизи продолговатого мозга, узла колена лицевого нерва или большого поверхностного каменистого нерва не влечет за -собой изменений в величине общего кровяного давления. Кроме того, расширение артерий наблюдается только на стороне раздражаемого нерва. В то же время перерезка большого поверхностного каменистого нерва предотвращает расширение артерий при раздражении лицевого нерва.
Все это вместе взятое может указывать на то, что импульс, идущий по пути, который указали Кобб и Хоробский, действительно представляет собой сосудорасширяющее влияние, изменяющее состояние артериальной стенки при непосредственном действии на нее. Эфферентная часть дуги этого рефлекса идет из области расположения ядра лицевого нерва в продолговатом мозгу к коленчатому ганглию. Затем по большому поверхностному каменистому нерву сосудорасширяющий импульс доходит до сплетения внутренней сонной артерии и направляется к сосу-
дам мозга. Раздражение по ходу указанного пути имеет своим следствием расширение артерий мягкой мозговой оболочки теменной области на 18% от исходного диаметра их (рис. 132).
Выше указывалось, что при раздражении вестибулярного аппарата мы установили расширение артерий мягкой мозговой оболочки лобно-теменной области (Б. Н. Клосовский, 1942). Рецепторным полем данного рефлекса являлся вестибулярный аппарат, а конечным полем рефлекса — артерии лобно-теменной области. Специфичность раздражения вестибулярного аппарата именно для сосудов указанных областей мягкой мозговой оболочки отчетливо выявлялась при одновременном наблюдении через «окно» сосудов лобной и затылочной области. Расширению артерий в первой из названных областей на 50% по сравнению с величиной их до опыта сопутствовало некоторое сужение сосудов в затылочной области.
Рефлекторный характер реакции артерий мягкой мозговой оболочки при раздражении вестибулярного аппарата находил свое подтверждение в том, что расширение во всех без исключения опытах продолжалось точно в течение 11/2 минут.
Увеличение диаметра артерий не могло быть отнесено за счет падения общего кровяного давления, поскольку падение общего кровяного давления обычно ведет к расширению артерий во всех без исключения областях мозга. В наших же экспериментах расширение артерий отмечалось лишь в определенной области мягкой мозговой оболочки, тогда как в других областях ее наблюдалось уменьшение просвета артерий. Специальные опыты, предпринятые нами совместно с Семеновым, показали, что падение кровяного давления, наблюдавшееся при раздражении вестибулярного аппарата, незначительно и непродолжительно. Кровяное давление обычно падало на 20—30 мм и удерживалось на этом уровне в течение лишь 15—30 секунд (Б. Н. Клосовский и В. Н. Семенов, 1947).
На следующем этапе разработки данного вопроса мы поставили задачу — установить промежуточные звенья по ходу сосудорасширяющего импульса от вестибулярного аппарата к сосудам лобной доли. В этом отношении большое значение имели полученные в наших прежних работах данные, на основании которых мы получили возможность составите схему хода вестибулярных импульсов для передачи на вегетативные центры (Б. Н. Клосовский, 1939). Как было нами установлено в соответствии с некоторыми уже имеющимися данными, импульсы, возникающие при раздражении вестибулярного аппарата, идут в триангулярное ядро, часть которого несет вегетативные функции. Отсюда импульсы могут следовать, с одной стороны, в ретикулярную субстанцию продолговатого мозга непосредственно к располагающимся там центрам, ведающим регуляцией общего кровяного давления. С другой стороны, по перекрещенной вестибуло-мезэнцефалической системе импульсы достигают среднего мозга и оканчиваются частью в области расположения ядер глазодвигательного нерва и частью в зрительном бугре.
Прилагаемая схема (рис. 120) может служить иллюстрацией, позволяющей с большей отчетливостью представить путь вестибулярного импульса к среднему мозгу.
С целью установления места переключения вестибулярного импульса на преганглионарный нейрон двигательной части рефлекторной дуги мы предприняли ряд экспериментов с изолированной перерезкой вестибуло-мезэнцефалических путей около сильвиева водопровода на дне четвертого желудочка.
Опыты, проведенные па кроликах, заключались в следующем: животному под уретановым наркозом в передней части черепа над лобно-те-менной долей ввинчивалась линза от микроскопа в металлической оправе. В образовавшееся таким образом окне в герметически закрытом черепе посредством капилляроскопа можно было наблюдать сосуды мягкой мозговой оболочки соответствующей области.
|
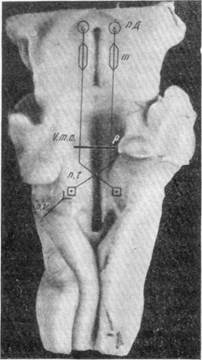
На следующем этапе эксперимента вскрывались кости и открывалась задне-черепная ямка. Шпателем осторожно приподнимался мозжечок над ромбовидной ямкой. Затем с помощью специально приготовленного бульботома поперечным движением производилась перерезка заднего продольного пучка, в состав которого входят пути от триангуляторного ядра к ядрам среднего мозга и зрительному бугру (место разреза заднего продольного пучка — Р показано на рис. 136). Раздражение вестибулярного нерва калорическим путем после перерезки заднего продольного пучка на дне ромбовидной ямки приводило к понижению общего кровяного давления, но расширение сосудов лобной доли отсутствовало.
| Рис. 136. Ромбовидная ямка кролика с проекцией проводящих путей от вестибулярных ядер к среднему мозгу. Р — место перерезки этих путей. |
Следовательно, можно было заключить, что переход вестибулярного импульса на преганглионарный нейрон двигательной части рефлекторной дуги осуществляется в районе среднего мозга, тогда как переход импульса к сосу-додвигательным центрам происходит в продолговатом мозгу. Таким образом, район поисков центра сосудодвигатель-ного рефлекса для сосудов головного мозга был ограничен средним мозгом, а путь, по которому идет этот рефлекс,— задним продольным пучком. Отсюда, естественно, встал вопрос, какому из ядерных образований среднего мозга принадлежит роль вазодилятаторного центра, регулирующего мозговое кровообращение.
С целью выяснения, какое ядро серого вещества, окружающего сильви-
ев водопровод, является источником преганглионарных волокон сосудо-расширителей, мы поставили опыты раздражения этих ядер электрическим током.
В настоящее время к вегетативным ядрам среднего мозга относят ядро n. supratrochlearis, ядро Якубовича-Вестфаль-Эдингера, ядро Даркшевича.
Первое из названных ядер, которое различные авторы именуют различно, располагается в сером веществе, окружающем сильвиев водопровод, над ядром блокового нерва. Ядро это резко отличается от лежащего рядом блокового нерва малым количеством тонких миэлиновых волокон и многочисленными безмиэлиновыми волокнами.
Парное ядро Якубовича-Вестфаль-Эдингера, состоящее из мелких, плотно расположенных клеток, лежит в окружении миэлиновых волокон мелкого калибра. Само ядро, по данным Пахе (Pache, 1935), бедно миэ-линовыми волокнами. Наряду с тонкими миэлинизированными волокнами, расположенными в правильном порядке, в ядре можно найти довольно толстые волокна неизвестного назначения, а также голые осевые цилиндры. Ядро, являясь вегетативно-моторным по своей природе, играет, по мнению большинства исследователей, решающую роль в иннервации гладкой мускулатуры глаза.
Ядро Даркшевича лежит на уровне задней спайки мозга у места перехода сильвиева водопровода в третий желудочек. Оно расположено в сером веществе и окружено восходящими волокнами задней спайки мозга, начало которых и следует, по мнению изучавших его авторов, отнести к данному ядру. Большое количество миэлиновых волокон, обнаруживаемых в области расположения ядра Даркшевича, обычно весьма незначительно по своему калибру.
Функции перечисленных ядер мало изучены. Так, например, Кахаль высказал предположение о происхождении волокон задней спайки из клеток верхнего двухолмия. Таким образом, направление аксонов ядра Даркшевича должно быть иное и поэтому даже по связям ядра с другими ядерными образованиями нельзя сделать предположения о возможной функции его. Невольное сомнение возникает также в отношении ядра Якубович-Вестфаль-Эдингера. Трудно представить себе, что такое большое количество нервных клеток служит только для снабжения двух не больших мышц глаза: круговой и ресничной.
На этих ядрах мы и сосредоточили внимание при попытке обнаружить в среднем мозгу центр сосудорасширяющей регуляции мозгового кровообращения.
Предпринятая в этом направлении работа была выполнена нами совместно с Е. Н. Космарской и М. Е. Афанасьевым. Материалом для исследования служили различные животные, в основном кролики.
На первом этапе эксперимента животному под наркозом вставлялось окно в лобно-теменную область. Через окно в течение всего опыта производилось наблюдение сосудов мягкой мозговой оболочки, измерение их и фотографирование.
В дальнейшем открывалась задняя черепная ямка. Для того чтобы избегнуть возможности раздражения самой ромбовидной ямки механическим или электрическим путем в ходе опыта, мы избрали разработанный нами подход к «раструбу» сильвиева водопровода.
Подход к «раструбу» сильвиева водопровода заключался в следующем. После вскрытия твердой мозговой оболочки и обнаружения червя мозжечка мы коагулировали располагавшиеся на нем сосуды. Главным образом коагулировались вены, особенно те из них, которые впадают в синусы твердой мозговой оболочки. После этого легко обнажались верхние края заднего двухолмия. Ложечкой убиралось вещество заднего червя мозжечка, прикрывающего вход в сильвиев водопровод. Вследствие предварительно произведенного коагулирования сосудов на поверхности червя мозжечка эта манипуляция проходила без кровотечения. При таком подходе к «раструбу» сильвиева водопровода вся ромбовидная ямка оставалась закрытой основной массой червя мозжечка и была защищена от какого-либо повреждения или раздражения.
Медные электроды, изолированные, за исключением концов их, бакелитовым лаком, перед введением в базальную область сильвиева водопровода укреплялись в специальном приборе. С помощью кремальер
"272
они могли быть продвинуты в любом направлении и на желаемую глубину. Ток для раздражения обычно брался от индукционной катушки. Сила раздражения составляла 15 см. Общий вид опыта представлен на рис. 137.
При постановке опыта вначале ток включался только тогда, когда электроды вводились до места расположения вегетативных ядер, лежащих над ядром блоковидного нерва. Затем электроды продвигались дальше и ток включался, когда электроды должны были находиться в области ядер Якубович-Вестфаль-Эдингера. Третий раз ток включался, когда электроды находились в области ядер Даркшевича.
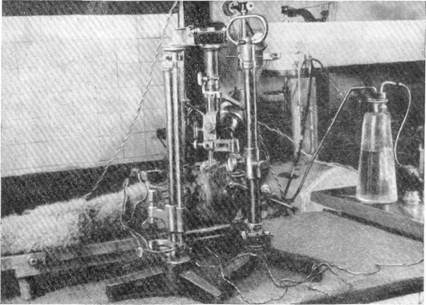
Рис. 137. Общий вид опыта с раздражением ядер Якубович-Вестфаль-Эдингера
и Даркшевича.
В дальнейшем, когда мы убедились, что раздражение вегетативного ядра блоковидного нерва не ведет к изменению просвета артерий мягкой мозговой оболочки, мы сразу вводили электроды до ядер Якубович-Вестфаль-Эдингера и Даркшевича. В конце опыта положение концов электродов маркировалось отложением меди в ткани с концов электродов после пропускания постоянного электрического тока. Затем средний мозг заливался в парафин и резался на серии срезов толщиной по 15 м. Срезы обрабатывались гематоксилин-эозином или по методу Ниссля.
Из 17 опытов на кроликах в 13 случаях наблюдалось вполне отчетливое расширение сосудов мягкой мозговой оболочки в лобно-теменной области. Это расширение, прослеженное нами на артериях калибром в 40—60 м, сочеталось с сужением зрачка и некоторым движением глаза вперед и кнутри.
При наблюдении сосудов соответствующей области через капилля-роскоп было отмечено, что между началом раздражения ядер в базаль-ной области сильвиева водопровода и моментом, с которого начинается расширение артерии, протекает краткий период. После определенного
латентного периода имеет место постепенное расширение артерий, сопровождающееся увеличением скорости тока крови в ней. Эти два явления оканчивались через 30 секунд.
Диаметр артерий во время раздражения обычно превышал просвет ее в норме на 40—50%.
Вслед за прекращением раздражения артерии в течение некоторого времени сохранили свой широкий просвет. Затем диаметр их постепенно уменьшался и артерии возвращались к своему исходному состоянию.
Дата добавления: 2015-05-16; просмотров: 1105;