ВОЗДЕЙСТВИЯ 3 страница
Макроскопический просмотр области раздражения во всех случаях
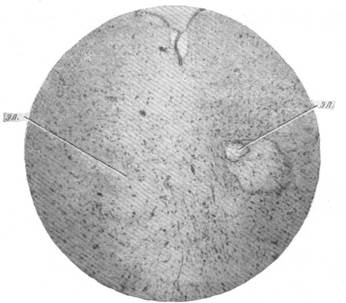
Рис. 138. Микрофотография, показывающая посредством маркировки расположение раздражающих
концов электродов (эл) в ядре Даркшевича.
Окраска по методу Ниссля. Увеличение 50.
показал, что электроды находились в базальной части серого вещества, окружающего сильвиев водопровод, в области переднего двухолмия. Иначе говоря, раздражению подвергалась область расположения вегетативных ядер Даркшевича и Якубович-Вестфаль-Эдингера.
В 6 случаях было произведено гистологическое исследование среднего мозга для уточнения места нахождения раздражавших электродов. Из них положительный результат в отношении раздражения ядер Дарк-шевича и Якубович-Вестфаль-Эдингера был получен в 5 опытах. В одном — гистологическое обследование дало отрицательный результат.
В положительных опытах раздражающие концы электродов находились или в самих ядрах Даркшевича, или в соприкосновении с ними (рис. 138). При отрицательном эффекте, полученном в одном из случаев, наблюдалось почти полное разрушение ядра Даркшевича, один из электродов при этом находился в ядре Якубович-Вестфаль-Эдингера.
Полученные факты дают нам право заключить, что расширение артерий мягкой мозговой оболочки связано с раздражением ядра Дарк-
шевича. Возникает, однако, вопрос, насколько возможно исключить в этих случаях влияние со стороны ядра Якубович-Вестфаль-Эдингера.
|
В проделанных экспериментах, как уже указывалось, расширение артерий постоянно сопровождалось реакцией со стороны зрачка и некоторым движением глазного яблока. Биполярное раздражение всегда имеет следствием рассеивание тока на некоторую область, располагающуюся вне пределов зоны раздражения. Отсюда можно думать, что реакции, сопутствовавшие расширению артерий, являются следствием раздражения части ядра Якубович-Вестфаль-Эдингера петлями тока.
Однако вопрос об участии ядра Якубович-Вестфаль-Эдингера в реакции расширения артерий лобно-теменной области может быть окончательно решен только пос-ле постановки опытов с перерезкой глазодвигательного нерва и с наблюдением процессов дегенерации в ядрах Даркшеви-ча и Якубович-Вестфаль-Эдингера. Ведущиеся в этом направлении опыты дадут ответ на поставленный вопрос и представят возможность с большим основанием отнести расширение сосудов за счет того или другого ядра.
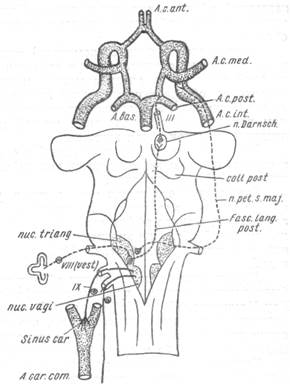
| Рис. 139. Схема парасимпатической иннервации сосудов мозга. |
В настоящее время полученные в наших опытах факты заставляют думать, что ядро Даркшеви-ча является источником преганглионарных волокон, идущих в составе глазодвигательного нерва. Волокна эти парасимпатической природы оканчиваются на разных уровнях своего хода на многочисленных нервных клетках, располагающихся в стенке сосудов. Второй нейрон, начинающийся от этих клеток, представляет собой конечный путь для сосудорасширяющего импульса.
Таким образом, подводя итоги всему сказанному по вопросу о характере влияния парасимпатической части вегетативной нервной системы на артерии мягкой мозговой оболочки, можно сделать следующие выводы.
1. В настоящее время можно считать установленным существование пути сосудорасширяющего импульса для артерий мягкой мозговой оболочки лобно-теменной области. Рецепторным полем этого рефлекса является вестибулярный аппарат, раздражение которого ведет к расширению артерий в указанной области на 50% по сравнению с исходной величиной их (рис. 139).
2. Установлена также часть пути сосудорасширяющего импульса для сосудов теменной доли, начало которого относится к области расположения ядра п. intermedius. Раздражение по ходу этого пути имеет своим следствием расширение артерий мягкой мозговой оболочки теменной доли на 18% от первоначального диаметра. Наличие периферического рецепторного поля для этого рефлекса, однако, окончательно не установлено. Не выяснено также, является ли раздражение большого поверхностного каменистого нерва специфическим только для артерий названной области, так как в работе Пенфильда (1932), посвященной этому вопросу, наблюдение сосудов производилось лишь в теменной области.
После рассмотрения современного состояния наших знаний по вопросу о влиянии симпатической и парасимпатической части вегетативной нервной системы на артерии мягкой мозговой оболочки остановимся коротко на обзоре существующих литературных данных относительно действия симпатико- и парасимпатикотропных веществ при непосредственном приложении их к артериям мягкой мозговой оболочки или при введении их в ток крови животного.
Известно, что артерии всех тканей и органов, за исключением легких и сердца, отчетливо сокращаются под действием адреналина даже в незначительной концентрации.
Существуют сообщения ряда авторов, указывающих на расширение артерий мягкой мозговой оболочки при локальном приложении к ним адреналина. Однако эффективность действия адреналина на сосуды мягкой мозговой оболочки значительно меньше, чем на сосуды других органов и тканей [Галлибуртон (Halliburton), 1910, Kay (Cow), 1911, и др.].
Другие исследователи пишут об отсутствии вообще каких бы то ни было реакций артерий при том же действии адреналина [Флори, 1925; Хау Маккинли (Howe McKinley), 1927; Мериль (Meriel), 1929, и т. д.] и, наконец, некоторые авторы считают, что артерии мягкой мозговой оболочки при местном приложении адреналина сокращаются (Форбс и Вольф, 1928; Форбс и сотр., 1933; Фог, 1939, и др.).
Такое расхождение результатов различных экспериментаторов может быть объяснено прежде всего незначительной величиной сокращения артерий при действии на них адреналина. Современный метод измерения величины артерий мягкой мозговой оболочки посредством окуляр-микрометра и специальных приборов через окно, вставленное в череп, показал, что в закрытом черепе артерии уменьшают свой просвет при действии на них адреналина всего на 4—8%. Такое незначительное изменение калибра артерий с трудом может быть отмечено при помощи специального прибора. Этим обстоятельством, вероятно, и объясняются противоречия в выводах исследователей, использовавших в своих работах простое наблюдение за сосудами мягкой мозговой оболочки в условиях открытого черепа.
Причиной может являться также то, что действие одной и той же концентрации сосудосуживающих веществ в зависимости от величины артерии проявляется различным образом. Так, например, согласно наблюдениям Фога (1939), местное приложение адреналина не вызывает реакций со стороны артерий, диаметр которых менее 100 м, тогда как артерии большего размера неизменно сокращаются.
Современные исследования указывают на сокращение артерий мягкой мозговой оболочки диаметром более 100 м при непосредственном действии на них сосудосуживающих веществ. Уменьшение просвета артерий
составляет 4—8% от исходной величины и значительно меньше того сужения артерий, которое наблюдается при действии тех же веществ в других тканях и органах [Фог, 1939; Луни и Фог (Lunn a. Fog), 1939, и т. д.].
Изучение непосредственного действия на артерии мягкой мозговой оболочки парасимпатикотропных веществ показало, что холиноподобные вещества вызывают расширение артерий более 30—100 м в диаметре [Финезингер, 1932; Беделл Томас (Bedell Thomas), 1937; Лунн и Фог, 1939]. Интересным оказывается действие кофеина. При наркотизации животного амиталом натрия непосредственное действие кофеина на артерии мягкой мозговой оболочки, по данным Финезингера, вызывает расширение артерий. Расширение артерий не сопровождается изменением общего кровяного давления.
При наркотизации животного эфиром местное приложение кофеина к артериям мягкой мозговой оболочки не оказывает влияния на просвет артерий, но при выходе животного из наркоза влечет за собой расширение артерий. Таким образом, результаты опытов Финезингера отчетливо указывают на зависимость реакций артерий мягкой мозговой оболочки при действии на них сосудорасширяющих веществ от характера наркоза и степени его.
В литературе имеются отдельные указания относительно действия растворов органических и неорганических веществ при непосредственном приложении их к стенке артерии мягкой мозговой оболочки. Так, например, 1% и 2% растворы солей стрихнина вызывают расширение артерий, азотнокислый натрий — слабое расширение, хлористый барий в определенной концентрации — сужение и т. д. (Флори, 1925, и др.).
Выяснение характера действия различных веществ непосредственно на стенку артерий мягкой мозговой оболочки имеет большое значение, так как знания по этому вопросу могут быть использованы во время операций на мозгу. Однако неточность и неопределенность результатов, полученных в эксперименте на животных, не позволяют пока использовать имеющиеся данные в практике нейрохирургических операций на человеке.
Выше указывалось, что артерии мягкой мозговой оболочки диаметром более 100 м испытывают непосредственное действие адреналина при местном приложении его к наружной стенке. Такие же соотношения наблюдаются и при введении адреналина в ток крови животного. Так, в опытах Фога (1939) было установлено, что в условиях предотвращения повышения общего кровяного давления (всегда имеющего место при введении в сосудистое русло адреналина) артерии диаметром более 100 м сокращаются на 4—8% по сравнению с величиной их до опыта. Сопоставление выше сказанного с результатами, полученными при раздражении симпатического нерва, дает возможность констатировать, что артерии мягкой мозговой оболочки находятся под незначительным влиянием симпатического отдела вегетативной нервной системы. Возбуждение последней имеет своим следствием сокращение указанных артерий на 4—8%.
Артерии, диаметр которых равен менее чем 50 м,не реагируют на непосредственное приложение сосудосуживающих веществ к наружной и внутренней стенке. Благодаря этому обстоятельству они могут быть использованы в качестве объекта в опыте, ставящем своей целью проследить характер ответных реакций артерий мягкой мозговой оболочки при введении в ток крови веществ, повышающих общее кровяное давление.
Предпринятые в этом направлении опыты показали, что повышение общего давления, вызванное введением в ток крови адреналина, раздражением чревного нерва или зажатием брюшной аорты, сопровождается сокращением артерий менее 50 м в диаметре. Размер сокращения зависит от уровня кровяного давления до опыта и от величины, на которую

|
оно возрастает (Фог, 1939; Форбс, Финлей, Нэсон, Ризе, 1929, 1933) (рисунок 140).
| Рис НО. Реакции артерий мягкой мозговой оболочки при повышении общего кровяного давления, вызванного введением в ток адреналина (по Фогу). На верхнем рисунке—запись реакции артерий при наличии более высокого уровня кровяного давления в начале опыта. Внизу—реакция артерий в условиях более низкого кровяного давления. |
При наличии давления, равного 110—140 мм, дальнейшее повышение давления дает лишь слабое сужение артерий мягкой мозговой оболочки, но артерии при этом не расширяются. Отсутствие расширения должно, повиди-мому, указывать на увеличение тонуса гладких мышечных волокон стенки артерий. Если же исходное давление ниже указанных цифр, то увеличение его на 20% имеет своим следствием сужение артерий мягкой мозговой оболочки. Сужение тем более выражено, чем ниже кровяное давление до опыта. В то же время з условиях очень низкого первоначального давления всякая регуляция тонуса артериальных стенок отсутствует. Есть также предел тонической регуляции и в другом отношении, а именно при внезапном резком увеличении
давления артерии не сокращаются, а расширяются.
Механизм сокращения артерий в ответ на повышение общего кровяного давления не установлен. Выяснено лишь, что данную реакцию можно считать независимой от влияния со стороны симпатического, блуждающего, синокаротидного и депрессорного нервов. Вполне возможно предположить, что повышение общего кровяного давления оказывает непосредственное влияние на тоническое состояние гладкой мускулатуры стенки артерий мягкой мозговой оболочки (Фог). Такое предположение мы считаем весьма вероятным.
Переходим к рассмотрению реакций артерий мягкой мозговой оболочки при различного рода состояниях асфиксии, вызванной замедлени-
ем скорости тока крови в сосудах мозга. Рассмотрим, таким образом, ответную реакцию артерий на низкое кровяное давление, на выключение сосудов, снабжающих головной мозг, а также на повышение внутричерепного давления.
Внезапное падение кровяного давления на значительную величину может быть получено при раздражении блуждающего, синокаротидного и депрессорного нервов, а также при раздражении поверхности каротид-ного синуса. Независимо от того, каким путем оно вызвано, падение общего кровяного давления имеет своим следствием расширение артерий мягкой мозговой оболочки. Полученные экспериментальным путем дан-
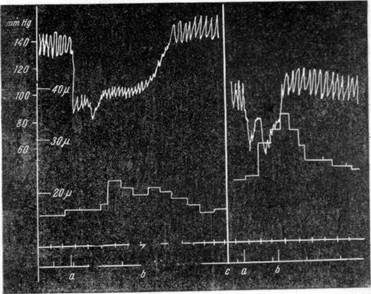
Рис. 141. Зависимость величины реакции артерий мягкой мозговой оболочки при падении общего кровяного давления от уровня его к моменту раздражения блуждающего нерва (а—b)
(по Фоту).
ные указывают, что величина, на которую при этом изменяется просвет артерий, в первую очередь зависит от уровня давления крови у животного к началу опыта.
Как можно видеть на рис. 141, большая сила реакции наблюдается в условиях низкого кровяного давления в начале эксперимента. Кроме того, величина изменения просвета артерий мягкой мозговой оболочки зависит также и от того, до какого уровня падает кровяное давление. На рис. 142 видно, что максимальное расширение артерий имеет место в случаях падения кровяного давления ниже 50 мм, т. е. ниже критического уровня его. Если же при значительном падении кровяного давления оно все же не опускается ниже 80 мм, со стороны артерий наблюдается только или незначительное расширение, или просвет их не изменяется.
Характерной особенностью поведения артерий мягкой мозговой оболочки при падении общего кровяного давления является сужение их, наблюдающееся тотчас же после начала снижения давления. Сужение
это кратковременно и продолжается 30—60 секунд. Сужение артерий с особой отчетливостью выступает при внезапном падении кровяного давления в какой-либо области мозга, вызванном закрытием снабжающего эту область сосуда. Так, в опытах нашей сотрудницы Е. Н. Космарской было отмечено, что закрытие средней или передней мозговых артерий постоянно сопровождается резким сужением артерий мягкой мозговой оболочки в областях выключенных сосудов. Вследствие сужения артерий мозговое вещество в указанных областях становилось бледным,
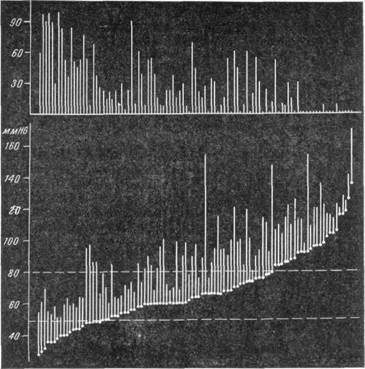
Рис. 142. Схема, показывающая зависимость изменения просвета артерий мягкой мозговой оболочки от исходного уровня кровяного давления и уровня, до которого оно снижается (по Форбсу).
В нижней части схемы представлены первоначальный уровень и
уровень, до которого падает кровяное давление. В верхней части—
соответствующее каждому случаю изменение диаметра артерий.
пульсация мозга прекращалась, скорость движения крови по сосудам резко замедлялась. Отчетливое сужение, наиболее выраженное у артерий среднего и особенно малого калибра, продолжалось в течение 30—120 секунд и не распространялось на вены, которые были заполнены кровью вплоть до наиболее мелких их ветвей.
Расширение артерий мягкой мозговой оболочки наблюдается не только при внезапном резком падении давления, но и при медленной кровопотере. На рис. 143 можно проследить, что и в этом случае падение давления сопровождается кратковременным сужением артерий, за которым следует расширение их (Фог, 1937; Б. Н. Клосовский, Е. Н. Космарская, 1950).
При изучении реакций артерий мягкой мозговой оболочки в ответ на постепенное снижение кровяного давления мы в совместных опытах с Е. Н. Космарской поставили своей целью проследить поведение не только артерий, но и вен в течение всего процесса медленной анемизации мозга.
На прилагаемых микрофотографиях (рис. 144, а, б, в, г, д, е, ж,з), взятых в качестве примера из нашей работы, обращает на себя внимание различный характер поведения артерий и вен при медленно развивающейся анемии головного мозга.
На микрофотографии (рис. 144, а) представлен, так сказать, исходный фон опыта. Как можно видеть, в поле зрения располагается артерия диаметром 58 ми три отходящие от нее веши диаметром 15, 40 и 32 м.
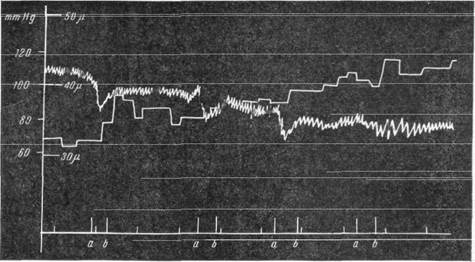
Рис. 143. Реакция артерий мягкой мозговой оболочки при постепенном выпускании
крови (по Фогу). В каждом из интервалов от а до b выпускалось 15 см3 крови.
Плотно заполненные кровью артерии располагаются на фоне не менее хорошо заполненных вен, диаметр которых равен 76, 63, 58 м.
Следующий снимок (рис, 144, б), сделанный через одну минуту после начала выпускания крови, дает возможность отметить изменение калибров артерий и вен, происходящее в противоположном направлении. Как можно видеть, артерии всех калибров резко расширились, но в то же время вены не менее заметно сократились.
При последующем выпускании артериальной крови увеличение диаметра артерий становится еще более значительно выраженным, но, наряду с расширением, намечаются первые признаки менее плотного заполнения артерий кровью.
В боковых ветвях основной артерии, а также и в венах красные кровяные шарики скапливаются большими или меньшими кучками, разделенными между особой плазмой. Вены по сравнению с предыдущей стадией несколько увеличивают свой размер, но остаются более узкими по сравнению с тем, какими они были в начале опыта.
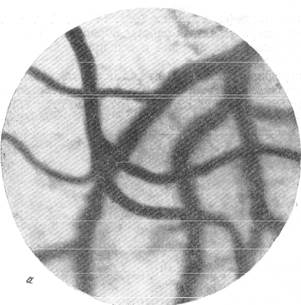
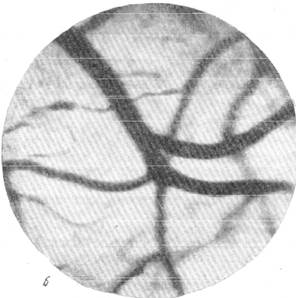
Рис. 144. Реакции артерий и вен мягкой мозговой оболочки кошки при постепенном
выпускании крови. а — спокойный сон; б — через 1 минуту после начала выпускания крови.
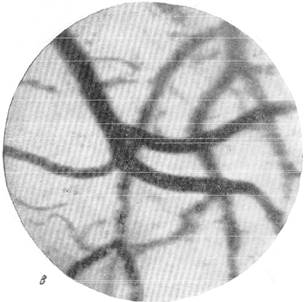

Рис. 144. Реакции артерий и вен мягкой мозговой оболочки кошки при постепенном
выпускании крови.
в — через 3 минуты после начала выпускания крови; г — через 25 секунд после
остановки дыхания.

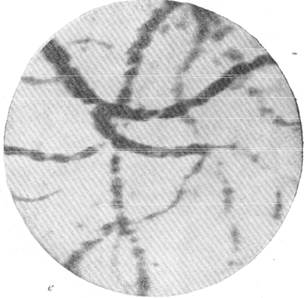
Рис. 144, Реакции артерий и вен мягкой мозговой оболочки кошки при постепенном
выпускании крови.
д — через 45 секунд после остановки дыхания; е — через 1 минуту 40 секунд после
остановки дыхания.
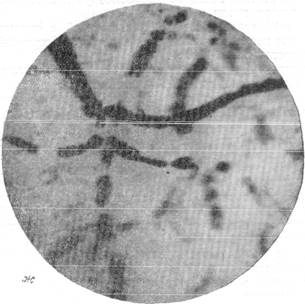
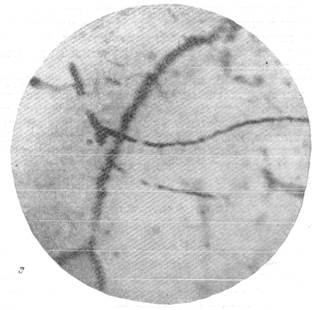
Рис. 144. Реакции артерий и вен мягкой мозговой оболочки кошки при постепенном
выпускании крови.
ж — через 2 минуты 30 секунд после остановки дыхания; з — через 12 минут после
остановки дыхания. Фото через «окно» в черепе с помощью капилляроскопа. Увеличение 60.
После остановки дыхательной деятельности артерии и вены в течение некоторого времени сохраняют размеры, которые они имели к моменту остановки дыхания (рис. 144. г, д, е). Изменение состояния сосудистой сети выражается лишь во вое более усиливающейся фрагментации в расположения крови в сосудах.
На следующей стадии, однако, наступившей в описываемом опыте через 2'/2 минуты после остановки дыхания, размер артерий уменьшился почти до исходного (рис. 144, ж).
Неравномерность заполнения артерий кровью стала еще более выраженной. При наблюдении в капилляроскоп перемещение крови по артериям и венам продолжалось еще довольно значительное время после остановки дыхательной и сердечной деятельности и прекратилось лишь через 12 минут.
На микрофотографии (рис. 144, з), снятой с мягкой мозговой оболочки уже после прекращения передвижения крови по сосудам, можно видеть, насколько резко выраженные изменения наступили в картине расположения сосудов.
Резко суженная артерия содержит кровь лишь в отдельных участках. Одна из ее ветвей не содержит крови совершенно. Вены сужены. Сосудистый рисунок стал очень бледным вследствие отсутствия крови во многих сосудах.
Проделанные эксперименты указывают, что кратковременное сужение артерий, наблюдающееся вслед за начинающим падать давлением, следует рассматривать как результат пассивного следования стенок сосуда за уменьшающимся объемом крови в нем. Наступающее в дальнейшем расширение артерий, по всей вероятности, связано с особенностями реакции со стороны гладких мышечных волокон сосудистой стенки в ответ на увеличивающееся содержание углекислоты в ткани мозга и в крови. Благодаря незначительному количеству мышечных волокон в стенке вен действие углекислоты на них крайне незначительно. Поэтому основным фактором, влияющим на просвет все, оказывается постепенно убывающее количество крови, поступающей к мозгу под все уменьшающимся давлением.
Анемия головного мозга, вызванная раздражением блуждающего и других нервов или выпусканием крови, постоянно сопровождается падением общего кровяного давления. Снизившееся давление или удерживается на низком уровне в течение всего времени раздражения, или постепенно понижается, как это имеет место при геморраялиях. При тако-го рода анемии головного мозга артерии или сразу расширяются до наибольшей величины в случаях резкого внезапного падания давления, или расширяются постепенно, по мере уменьшения крови в организме при потере ее. Расширившиеся артерии и суженные вены сохраняют свой просвет в продолжение всего времени, пока кровяное давление держится на низком уровне, и возвращаются к исходному размеру при повышении давления.
Несколько иная картина наблюдается при прекращении кровотока по основным артериям, снабжающим головной мозг. Сотрудница нашей лаборатории Е. Н. Космарская показала, что при выключении сонных и позвоночных артерий давление в сосудах виллизиева круга и в сосудах головного мозга резко падает и остается на низком уровне в продолжение 10—20 минут после зажатия сосудов. Общее давление при этом не только не падает, но, благодаря регуляторной деятельности каротидного синуса, значительно повышается Уровень повышения общего давления и различная у животных степень развития коллатеральных сосудов опре-
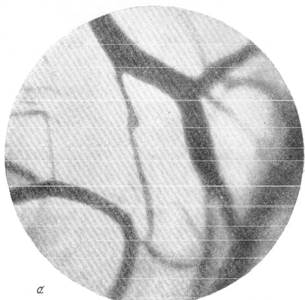
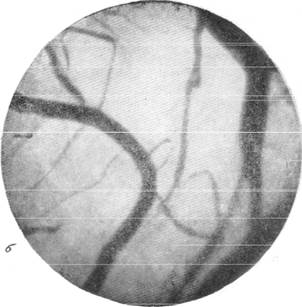
Рис. 145. Реакции сосудов мягкой мозговой оболочки кролика при зажатии обеих общих сонных артерий на шее (опыт без предварительной наркотизации животного), а — просвет артерий и вен до опыта; б — артерии и вены через 30 секунд после
зажатия сонных артерий.
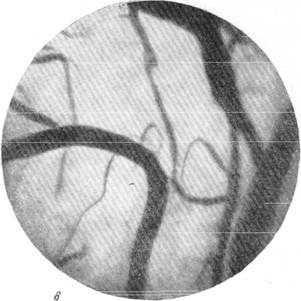
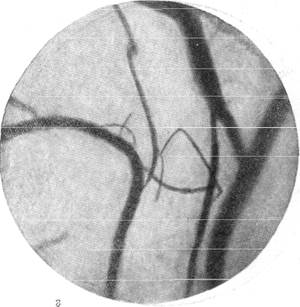
Рис. 145. Реакции сосудов мягкой мозговой оболочки кролика при зажатии обеих
общих сонных артерий на шее (опыт без предварительной наркотизации животного).
в — то же через l1/2 минуты; г—через 1 минуту после снятия зажимов с сонных артерий.
Фото через «окно» в черепе. Увеличение 60.
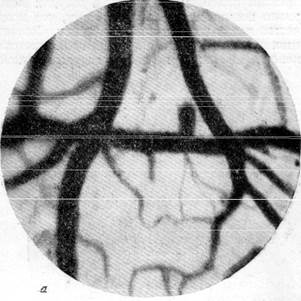
|
|
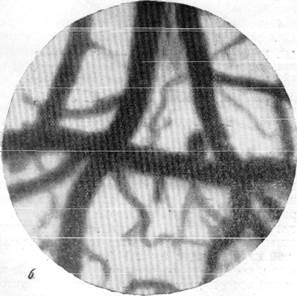
Рис. 146. Реакции артерий и вен мягкой мозговой оболочки кошки при зажатии обеих общих сонных и одной из позвоночных артерий на шее (опыт в условиях предварительной наркотизации).
а — спокойный сон; б—артерии и вены через 1 минуту 25 секунд после зажатия трех
артерий.
деляют скорость переключения мозга на кровоснабжение по коллатеральным сосудам, а тем самым лучшее или худшее питание его.
Таким образом, в случаях зажатия основных питающих мозг артерий вслед за резким падением давления в сосудах мозга оно по степенно и непрерывно повышается благодаря тому, что мозг получает все большее количество крови под высоким давлением. Соответственно с этим при выключении снабжающих мозг артерий наблюдается некоторое своеобразное изменение ширины просвета сосудов мягкой мозговой оболочки.
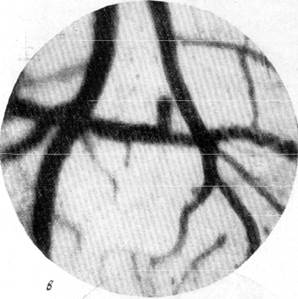
Рис. 146. Реакции артерий и вен мягкой мозговой оболочки кошки при зажатии обеих общих сонных и одной из позвоночных артерий на шее (опыт в условиях предварительной наркотизации).
в —через 1 минуту 10 секунд после снятия зажимов
с артерий. Фото через «окно» в черепе. Увеличение 60.
На микрофотографиях (рис. 145, а, б, в, г), взятых из работы, проделанной нами совместно с Е. Н. Космарской, представлены результаты одного из опытов на ненаркотизированном кролике. Можно видеть, что зажатие обоих общих сонных артерий на шее кролика без наркоза имеет своим следствием расширение не только артериальной,но и венозной части сосудистой сети мягкой мозговой оболочки. Те же соотношения мы наблюдаем у животных, у которых закрытие артерий производилось после предварительной наркотизации (рис. 146, а, 6, в).
Падение давления в сосудах головного мозга, следующее за выключением снабжающих его артерий, ведет к накоплению продуктов обмена веществ нервной ткани. Увеличенное содержание углекислоты вызывает расширение артерий мягкой мозговой оболочки. Уменьшение скорости тока крови сопровождается замедленным оттоком ее из вен, в результате чего вены также расширяются. По истечении некоторого времени с момента закрытия артерий и с увеличением поступления крови к мозгу
по коллатеральным сосудам просвет артерий и вей начинает уменьшаться. Освобождение артерий от зажимов ведет к быстрому возвращению диаметров артерий и вен мягкой мозговой оболочки к исходной величине.
Таким образом, можно сделать заключение, что замедление скорости тока крови и увеличение содержания углекислоты в ткани мозга являются непосредственной причиной расслабления самой сосудистой стенки.
Важнейшую роль в регуляции просвета артерий мягкой мозговой оболочки играет химический состав крови животного. Избыточное содержание в крови кислорода сопровождается уменьшением диаметра артерий, тогда как увеличение напряжения углекислоты влечет за собой расширение их.
Зажатие дыхательного горла у животного при нормальном поступлении крови к мозгу по интактным сонным и позвоночным артериям ведет к резкому расширению артерий и вен всех калибров мягкой мозговой оболочки (Б. Н. Клосовский, Е. Н. Космарская, 1950). Это ясно видно на микрофотографии (рис, 147, а, б, в, г), снятой с мягкой мозговой оболочки кошки, на дыхательное горло которой был наложен зажим. Прослеживая поведение сосудов мягкой мозговой оболочки в течение длительного времени, мы могли убедиться в чрезвычайнойсиле асфиксии как сосудорасширяющего фактора. В эксперименте, представленном на микрофотографиях (рис. 147), артерии и вены продолжали оставаться расширенными даже спустя более 20 минут после снятия зажима с трахеи. Более того, спустя более значительный интервал времени, когда артерии уже возвращались к исходной величине, вены оказывались еще заметно расширенными.
Таким образом, одновременное действие все увеличивающейся концентрации углекислоты на внутреннюю, обращенную к току крови, и на наружную ставку артерии ведет к расширению ее. Расширение тем более выражено, чем сильнее и продолжительнее асфиксия. Резкое увеличение общего кровяного давления, наблюдающееся в условиях асфиксии всего организма, имеет своим следствием увеличение тока крови по расширившимся артериям мягкой мозговой оболочки и пассивное растяжение вен,
Следовательно, анемическая или аноксическая асфиксия мозга ведет к расширению артерий мягкой мозговой оболочки. Расширение артерий является результатом изменении тонического состояния их стенки,, возникающего при непосредственном воздействии углекислоты на гладкие мышечные волокна, входящие в состав артериальной стенки. Иначе говоря, угольная кислота представляет собой мощный сосудорасширяющий фактор, играющий важнейшую роль в регуляции просвета сосудов мягкой мозговой оболочки.
По данным отдельных авторов, артерии мягкой мозговой оболочки испытывают регулирующее влияние и со стороны осмотического давле-ния находящейся в них крови (Форбс, Вольф, 1928). В то время как введение в ток крови животного изотонического раствора поваренной соли не вызывает никаких реакций со стороны артерий мягкой мозговой оболочки, введение в ток ирови или в брюшную полость гипертонических растворов ведет к сокращению их.
Однако такого рода реакция наблюдается не во всех случаях. Имеются сообщения, согласно которым введение гипертонических растворов не влечет за собой изменения просвета артерий мягкой мозговой оболочки [Хау и Мак Кинли, 1927). Таким образом, вопрос о влиянии
Дата добавления: 2015-05-16; просмотров: 1085;