СООТНОШЕНИЯМЕЖДУ НЕРВНЫМИ КЛЕТКАМИ И КАПИЛЛЯРАМИ
Интимное взаимоотношение нервных клеток и капилляров представ-ляет собой наиболее важный и интересный вопрос проблемы циркуляции крови в мозгу. До настоящего времени исчерпывающего ответа на этот вопрос не существовало, вследствие отсутствия методов, допускающих одновременное изучение клеточного я сосудистого строения головного мозга. Для того чтобы понять характер взаимодействия нервной клетки с окружающими ее капиллярами, необходимо получить ва одном и том же препарате оба компонента нервной ткани.
Существующие методы гистологической обработки позволяют выявлять и изучать в деталях только нервную клетку или капилляр. Попытки дополнить один метод другим пока не дали вполне удовлетворительных результатов. В самом деле, изучение клеточного строения нервной ткани требует гистологических срезов, не превышающих 30 м, но на срезах указанной толщины утрачивается трехмерное расположение сосудистой сети. При выявлении сосудистой сети моага с наибольшей полнотой на препаратах толщиной в 150—300 м дополнительная окраска с целью получения нервных клеток делает невозможным дальнейшее исследование вследствие совершенной непрозрачности (препарата. Пригодными для изучения взаимоотношений клеток и капилляров являются препараты с полностью представленной сосудисто-капиллярной сетью, на которых иногда оказываются импрегнированными также отдельные нервные клетки или группы их.
Среди серий препаратов, обработанных предложенным нами методом импрегнации сосудистой стенки серебром, можно найти такие, в отдельных участках которых оказываются выявленными одиночные нервные клетки, а иногда даже группы клеток. Воспользовавшись такими препаратами, мы прежде всего обратили внимание на взаимодействие капилляров я больших пирамидных нелегок коры.
На рис. 156, а, б, в, г видна одна из таких клеток, располагающаяся в капиллярной петле. Рассмотрение этих микрофотографий позволяет отметить, что при фотографирований до известной степени искажается диаметр капилляров, лежащих в различных плоскостях. Кажется, что капилляры, находящиеся в фокусе, имеют больший диаметр по сравнен нию с капиллярами, расположенными вне фокуса. Кроме того, при фокус -ном фотографировании почти полиостью исчезает представление о пространственном расположении сосудисто-капиллярной сети в мозгу. Указанные недостатки фотографической документации побудили нас допол-
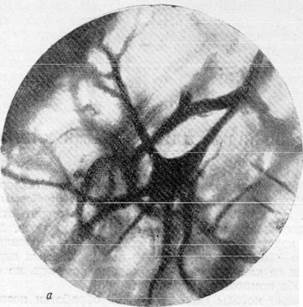

Рис. 156, а, 6. Микрофотографии, иллюстрирующие расположение пирамидных клеток
коры полушарий головного мозга в капиллярной петле. Импрегнация по методу Б. Н. Клосовского. Увеличение 400, 800.
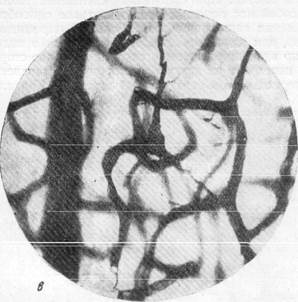
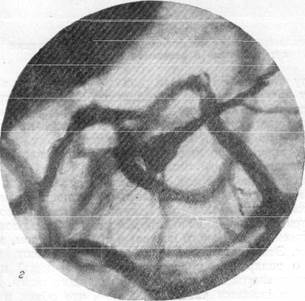
Рис. 156, в, г. Микрофотографии, иллюстрирующие расположение пирамидных клеток
коры полушарий головного мозга в капиллярной петле. Импрегнация по методу Б. Н. Клосовского. Увеличение 400 и 800.
нить и расширить ее с помощью рисунка. Мы зарисовывали пирамидную клетку со всеми ее отростками и окружающие клетку капилляры при каждом повороте микрометрического винта. При этом с возможной точностью отмечались диаметры капилляров и положение капилляров в отношении нервной клетки. При сравнении сделанных таким образом рисунков между собой нас поразило исключительное однообразие в от-ношении расположения капилляров возле большой пирамидной клетки. На рис. 157 видно, как капилляр огибает тело клетки в виде опирали. При этом капилляр вначале располагается возле одной из сторон клетки, затем, изгибаясь, переходит на другую ее сторону и оставляет тело нервной клеши при переходе ее в верхушечный отросток. Капилляр плотно

|
Рис. 157. Соотношение между большими пирамидными клетками артериальными и венозными капиллярами.
коры и
приложит к телу клетки. На том же рис. 157 видно, что в основании клетки проходит другой капилляр с несколько большим по сравнению с первым диаметром. Этот капилляр идет на некотором расстоянии от тела клетки, причем часть отростков последней находится Во одну сторону капилляра, часть — по другую. Благодаря характерному расположению этого капилляра, пирамидная клетка коры головного мозга как бы лежит своим основанием на этом капилляре.
Прослеживая дальнейший ход таких капилляров, мы в некоторых случаях могли ясно видеть, что капилляр, окружающий клетку в виде спирали, является артериальным, так как он представляет собой капиллярное продолжение артерии. Капилляр, лежащий в основании клетки, является продолжением венозного сосуда и, таким образом, может быть назван венозным. Эти морфологические данные делают возможным предположение о поляркости пирамидной клетки — одной из наиболее диференцированных клеток коры полушарий головного мозга. Полярность пирамидной клетки является результатом того, что обмен в ней имеет определенную направленность. Надо думать, что участии тела пирамидной клетки, прилежащие к артериальному капилляру, представляют собой место, где происходит интенсивное поглощение кислорода.
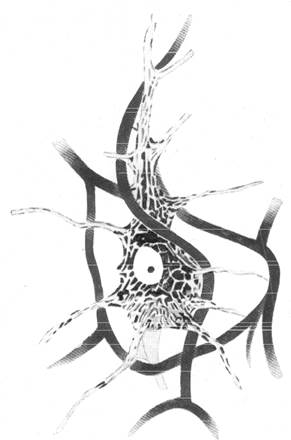
Рис. 158. Соотношение между распределением
нисслевской зернистости и пигмента в теле
большой пирамидной клетки коры и капиллярами
вокруг нее.
Напротив, в тех участках клетки, которые располагаются вблизи венозного капилляра, по всей вероятности, происходит отдача продуктов обмена веществ.
Если обратитьсяк особенностям тонного внутреннего строения большой пирамидной клетки коры больших полушарий, как оно выявляется на препаратах, окрашенных по методике Ниссля, то, согласно всем исследователям, пирамидная клетка асимметрична по своему (внутреннему строению. В качестве примера приводим рисунок (ряс. 158) большой пирамидной нервной клетки, взятый нами из руководства по неврологии с окружающими ее капиллярами, нанесенными на рисунок в соответствии с данными наших исследований. На этом рисунке видно, что нисслевская зернистость распределена в теле клетки неравномерно. Наибольшее скопление хроматиновых глыбок отмечается справа от ядра, где к тому же интенсивность окрашивания хроматиновых глыбок значительно выражена. Снизу от ядра вместо хроматина отмечается скопление пигмента липофусцина. В месте отхождения от клетки аксона конусообразное расширение тела клетки совершенно лишено и хроматиновых глыбок, и пигмента.
При сравнении расположения тигроида и липофусцина в большой пирамидной нервной клетке коры с расположением капилляров вокруг нее (рис. 157 и 158) можно видеть, что наибольшее количество нисслев-ской зернистости будет наблюдаться в местах клеток, прилежащих к артериальному капилляру.
Ближе к основанию пирамидной клетки, где сосредоточивается пигмент липофусцин, атакже находится большой участок, не содержащий ни пигмента, ни нисслевской зернистости (место отхождения аксона), проходит венозный капилляр.
Таким образом, тонкая структура пирамидной клетки коры указывает на определенную направленность процессов обмена веществ в ней. В соответствии с расположением артериального и венозного капилляров возле теша пирамидной клетки нужно думать, что обмен веществ происходит в направлении от верхушечного отростка к основанию большой пирамидной клетки коры полушарий головного мозга, т. е. в направлении импульса от дендритов к аксону. Вполне возможно предположить, что полярность и определенный способ взаимодействия с артериальной и венозной частью сосудистой сети мозгового вещества характерны только для высокодиференцированных больших пирамидных клеток.
На настоящем уровне наших знаний нельзя дать никакого определенного ответа на вопрос, существует ли полярность у тех нервных клеток, которые располагаются в одной капиллярнойпетле не в одиночку, а группами. По этому поводу можно высказать лишь общие соображения. Возможно, что и у этих клеток существует такая же полярность обмена веществ, как и у пирамидных клеток. Нет оснований, однако, отрицать наличие и другого способа обмена. Можно предполагать, что в определенный момент клетка только усваивает кислород и питательные вещества крови, тогда как в другой момент она только отдает продукты обмена веществ. Иначе говоря, в первом случае ассимиляция и диссимиляция протекают в клетке одновременно, во втором — в разное время.
Как известно, наиболее простые взаимоотношения между артериальной и венозной частью сосудистой сети существуют у тех животных, мозг которых получает кровоснабжение по анатомически конечным артериям. Мозг этих животных является чрезвычайно удобным объектом для выяснения способа взаимодействия капилляров с нервными клетками. Но решение данного вопроса возможно лишь в том случае, если
будет установлено взаимное расположение артерии и вены в пределах одной артерио-венозной единицы и расположение этих сосудов в различных единицах, К сожалению, несмотря на принципиальную важность доставленных вопросов, они не поднимались исследователями, изучавшими кровоснабжение мозга опоссума или кенгуру, и требуют дальнейшей экспериментальной разработке.Выяснение их безусловно поможет разобраться в тех сложных взаимоотношениях нервного и сосудистого компонентов нервной ткани, которые наблюдаются в мозгу более высоко организованных животных.
Важность выяснения соотношений между нервными клетками и капиллярами признают вое исследователи, занимающиеся изучением строения и функций головного мозга. Почти каждый исследователь, работающий с области кровоснабжения головного мозга, несмотря на отсутствие соответствующих методик для выявления соотношений между нервными клетками и капиллярами, считает необходимым тем или иным образом коснуться этого вопроса. Поэтому имеющиеся в литературе данные представляют собой соображения общего порядка, основывающиеся на разровненных фактах, позволяющих установить только главнейшие соотношения капиллярной сени мозга с нервными клетками.
Известно, что нора полушарий головного мозга имеет капиллярную сеть, сформированную из чрезвычайно плотно расположенных капиллярных петель. При сравнении приведенных микрофотографий (рис. 159, а, б) отчетливо выступает различие капиллярных сетей в сером и белом веществе и значительно лучшая васкуляризация первого по сравнению со вторым. Густота капиллярной сети в коре послужила поводом к предположению, согласно- которому в отдельных участках серого вещества капиллярные петли лежат так тесно друг к другу, что между ними могут пройти только отростки нервных клеток, но тела их поместиться не могут (Де Но, 1927). Однако последующие исследования показали, что, несмотря на густую капиллярную сеть, в одной капиллярной петле в мозгу животных, например, кошек, могут располагаться друг возле друга 10 и даже 16 нервных клеток. В то же время в мозгу могут быть обнаружены и такие участки, например, в locus eoeruleus, находящемся в верхне-боковой части ромбовидной ямки человека, где в значительной по размеру капиллярной петле лежит только одна нервная клетка (Пфайфер, 1928).
Какой же из двух этих основных компонентов мозга—нервная клетка или капилляр—определяет их взаимное расположение по отношению друг к другу? Рассмотрим первый ряд наблюдений, указывающих на зависимость. клеточного строения от характера кровоснабжения. В этом отношении интересны данные, сообщенные Кахалем (R. Kajai, 1929). Исследуя мозговые сосуды насекомых, Кахал нашел, что они окружены кровеносными сосудами, располагающимися по поверхности, но не проникающими вглубь ганглия. Соответственно этому нервные клетки в узле ле-жат поверхностно. Более того, ориентация нервных клеток к сосудам определяет и морфологическое оформление их, так как клетки в этих случаях униполярны. Подобные же клетки наблюдаются повсюду, где имеет место снабжение кровью только с периферии. По указанию Каха-ла, униполярный тип клеток преобладает в плотных нервных узлах не только у насекомых, но и у червей и моллюсков. Клетки того же характера составляют основную массу серого вещества шинного мозга ланцетника, лишенного внутренних кровеносных сосудов.
Иной тип нервных клеток встречается там, где имеет место проникновение сосудов с периферии внутрь нервных узлов. Так, например,
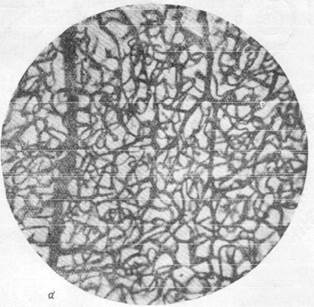
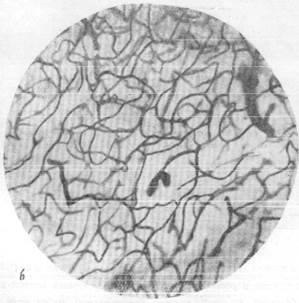
Рис. 159. Капиллярная сеть коры и белого вещества головного мозга кошки.
а — капиллярная сеть в сером веществе; б — капиллярная сеть в белом веществе.
Импрегнация по методу Б. Н. Клосовского. Увеличение 100.
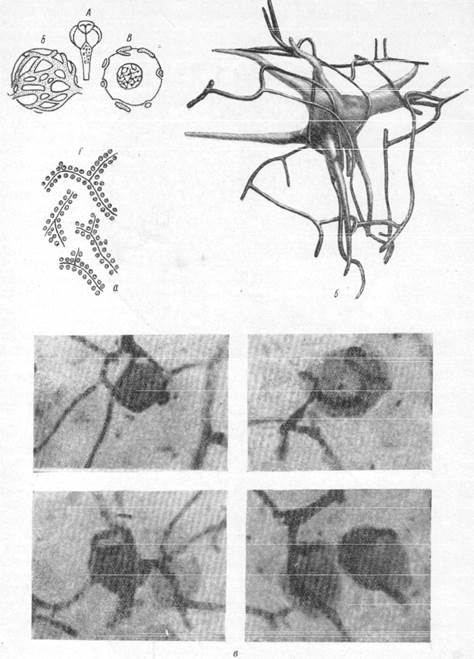
Рис. 160.
а — соотношение нервных клеток и капилляров: А — мозг акулы (Spheroides maculatus) с экстрамедуллярными нервными клетками; Б — сеть капилляров на экстрамедуллярной клетка; В — та же клетка в разрезе; Г — группировка нервных клеток вокруг сосудов в
зрительной доле каракатицы; б — соотношение большой нервной газетки ретикулярной субстанции с капиллярами
(реконструкция с препарата; увеличение 400):
в — микрофотографии, иллюстрирующие взаимоотношение нервных клеток и капилляров в мезэнцефалическом корешке тройничного нерва. Увеличение 480.
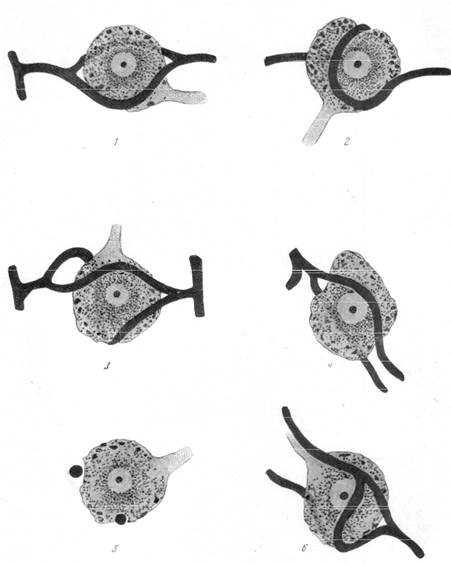
Рис. 160, г. Соотношение между нервными клетками и капиллярами в мезэнцефалическом корешке тройничного нерва.
в зрительном узле каракатицы, содержащем внутри большое количество сосудов и капилляров, клетки мультиполярны. Но и здесь можно проследить отчетливую зависимость расположения клеток от направления, в котором проходят кровеносные сосуды. Так, нервные клетки в зрительном узле каракатицы группируются вдоль разветвленийсосудистой системы (рис. 160, а—г). Сосуды представляют собой, образно говоря, оси, по обе стороны от которых лежат клетки. Количество клеток определяется диаметром сосуда. Возле центральных сосудистых стволов значительного диаметра клетки располагаются в несколько рядов, тогда как тонкие сосуды окружены одним рядом нервных клеток (Кахал, 1929; Шаррер, 1937).
Таким образом, в описанных случаях взаимное расположение клеток и сосудов определялось сосудами. Это указывает на стремление в определенных условиях нервных клеток к сосудам как к источникам питания.
Другие наблюдения показывают, что к капилляры стремятся к нервным клеткам. Подобные картины можно, например, наблюдать в некоторых вегетативных ядрах мозга, как nucleus magnooellularis praeoptieus в промежуточном мозгу рыб, и в гомологичных ядрах млекопитающих (Шаррер. 1937). Как видно на рис. 160, а, б и в, капилляры плотно прилежат к поверхности клетки, образуя как бы корзинку.
При изучении соотношения клеток и капилляров в ретикулярной субстанции продолговатого мозга сотрудники нашей лаборатории Е. Н. Космарская и Е. Г. Балашова (1950) обнаружили несколько различных способов взаимоотношений названных компонентов мозгового вещества в зависимости от величины нервных клеток. Работа велась методом прижизненной инъекции сосудистого русла трипановой синью с последующим докрашиванием нервных клеток крезилэхтвиолетом, а также нашим методом импрегнации сосудистой станки серебром.
На рис. 160, б представлена реконструкция большой нервной клетки моторного типа с расположенными возле нее капиллярами. Клетки, подобные изображенной на рис. 160, б, располагаются в ретикулярной субстанции продолговатого мозга, в области, которая то современным представлениям является местом локализации сосудодвигательного центра и имеет в длину 75 м и ширину — 35м. На рис. 160, б видно, что крупные клетки, входящие в состав сосудодвигательного центра, окружены большим количеством капилляров. Часть капилляров плотно прилежит к различным поверхностям тела клетки, проходя по ним в различных направлениях.
Тесные взаимоотношения с капиллярами обнаруживают не только тело, но и отростки больших клеток сосудодвигательного центра. Отростки в большинстве своем сопровождаются одним или чаще двумя-тремя Капиллярами, идущими вдоль отростка на некотором его протяжении. Часть отростков в месте отхождения от тела клетки оказывается обвитой капиллярами. Поскольку капилляры располагаются возле тел крупных клеток сосудодвигательного центра в различных плоскостях, фотография в значительной степени искажает действительное соотношение этих клеток с капиллярами. Поэтому при исследовании соотношения нервных клеток и капилляров Е. Н. Космарская и Е. Г. Балашева произвели реконструкцию такой клетки и окружающих ее капилляров. Вокруг клеток ретикулярной субстанции меньшего размера -от 50 до 30 м в длину - не отмечается такого большого количества капилляров, как возле описанных выше крупных клеток. Однако клетки длиной 50—30 м как в области сосудодвигательного, так и в области дыхательного центра всегда имеют один капилляр,
тесно прилежащий к какой-либо одной поверхности нервной клетки. Часть отростков нервных клеток указанных размеров в обоих центрах также на некотором своем протяжении сопровождается капиллярами. Наконец, наиболее мелкие по размерам клетки сосудодвигательного и дыхательного центров длиной 20—8 м не обнаруживают какого-либо отношения к капиллярам и располагаются свободно в крупных капиллярных петлях ретикулярной субстанции.
Контакт между нервной клеткой и капилляром может быть настолько тесным, что отдельные капилляры внедряются в протоплазму клетки и располагаются в бороздках или канавках, проходящих по поверхности тела клетки. Такого рода характерное взаиморасположение клеток и капилляров было обнаружено не только у рыб, но и у морских свинок и даже у человека. Коллин описал подобные перицеллюляр-ные капилляры в зубчатом ядре мозжечка и в вегетативных ядрах гипоталамуса. Сотрудница нашей лаборатории Е. Г. Балашева (1950) подробно проследила взаимоотношение между нервными клетками мезэнце-фалического корешка тройничного нерва и капиллярами. Характерной особенностью этих взаимоотношений является тесное прилежание капилляров к поверхности тела нервной клетки, имеющей шарообразную форму и один только отросток, а также расположение капилляров в желобке протоплазмы клетки (рис. 160, г, 5).
На микрофотографии (рис. 160, в) и на схеме (рис. 160, г) представлены разные типы взаимоотношений между нервной клеткой мезэнцефа-лического корешка и капиллярами. В ряде случаев к телу нервной клетки подходит артериальный капилляр, который, раздваиваясь, охватывает тело нервной клетки по экватору или же изгибается по двум сторонам тела клетки, затем снова соединяется в один уже венозный капилляр (рис. 160, г, 1,3,4,6). Такое разделение капилляра на два дугообразных с последующим .соединением снова в один мы неоднократно наблюдали на препаратах сосудистой сети, импрегнированной серебром по нашей методике, в коре больших полушарий и особенно в поле Брока.
В другом ряде случаев капилляр, подходя к телу клетки, охватывает его спирально, плотно прилегая к протоплазме клетки и даже частью вдавливаясь в нее (рис. 160, г, 2).
В литературе имеются указания о существовании нервных клеток, взаимодействие которых с питающим капилляром заходит так далеко, что капилляр проходит сквозь тело клетки и является, таким образом, внутриклеточным. Клетки с внутриклеточными капиллярами встречаются редко, они описаны у рыб, в поле 4 коры человека, а также в nucleus supraopticus и nucleus paraventricularis [Студничка (Studnicka), 1903; Бильшовский, 1928]. Своеобразное расположение капилляра внутри тела нервной клетки является, по мнению ряда авторов, отражением особой напряженности функциональной деятельности тех клеток, сквозь тело которых они проходят.
Мы не могли точно установить наличие внутриклеточных капилляров в центральной нервной системе и полагаем, что если они и существуют, то не могут являться результатом прорастания капилляра через тело нервной клетки. Проще представить себе, что на какой-то стадии развития капилляр располагается в непосредственной близости от поверхности тела нервной клетки, протоплазма которой при дальнейшем росте окружает капилляр со всех сторон и он, таким образом, оказывается включенным в нее.
Итак, мы видим, что в литературе существуют два взгляда на взаимоотношение нервных клеток и капилляров. Согласно первой точке зре-
тесно прилежащий к какой-либо одной поверхности нервной клетки. Часть отростков нервных клеток указанных размеров в обоих центрах также на некотором своем протяжении сопровождается капиллярами. Наконец,.наиболее мелкие по размерам клетки сосудодвигательного и дыхательного центров длиной 20—8 м не обнаруживают какого-либо отношения к капиллярам и располагаются свободно в крупных капиллярных петлях ретикулярной субстанции.
Контакт между нервной клеткой и капилляром может быть настолько тесным, что отдельные капилляры внедряются в протоплазму клетки и располагаются в бороздках или канавках, проходящих по по-верхности тела клетки. Такого рода характерное взаиморасположение клеток и капилляров было обнаружено не только у рыб, но и у морских свинок и даже у человека. Коллин описал подобные перицеллюляр-ные капилляры в зубчатом ядре мозжечка и в. вегетативных ядрах гипоталамуса. Сотрудница нашей лаборатории Е. Г. Балашева (1950) подробно проследила взаимоотношение между нервными клетками мезэнце-фалического корешка тройничного нерва и капиллярами. Характерной особенностью этих взаимоотношений является тесное прилежание капилляров к поверхности тела нервной клетки, имеющей шарообразную форму и один только отросток, а также расположение капилляров в желобке протоплазмы клетки (рис. 160, г, 5).
На микрофотографии (рис. 160, 0) и на схеме (рис. 160, г) представлены разные типы взаимоотношений между нервной клеткой мезэнцефа-лического корешка и капиллярами. В ряде случаев к телу нервной клетки подходит артериальный капилляр, который, раздваиваясь, охватывает тело нервной клетки по экватору или же изгибается по двум сторонам тела клетки, затем снова соединяется в один уже венозный капилляр (рис. 160, г, 1,3,4,6). Такое разделение капилляра на два дугообразных с последующим .соединением снова в один мы неоднократно наблюдали на препаратах сосудистой сети, импрегнированной серебром по нашей методике, в коре больших полушарий и особенно в поле Брока.
В другом раде случаев капилляр, подходя к телу клетки, охватывает его спирально, плотно прилегая к протоплазме клетки и даже частью вдавливаясь в нее (рис, 160. г, 2).
В литературе имеются указания о существовании нервных клеток, взаимодействие которых с питающим капилляром заходит так далеко, что капилляр проходит сквозь тело клетки и является, таким образом, внутриклеточным. Клетки с внутриклеточными капиллярами встречаются редко, они описаны у рыб, в поле 4 коры человека, а также в nucleus supraopticus и nucleus paraventricularis [Студничка (Studnicka), 1903; Вильнюсский, 1928]. Своеобразное расположение капилляра внутри тела нервной клетки является, по мнению ряда авторов, отражением особой напряженности функциональной деятельности тех клеток, сквозь тело которых они проходят.
Мы не могли точно установить наличие внутриклеточных капилляров в центральной нервной системе и полагаем, что если они и существуют, то не могут являться результатом прорастания капилляра через тело нервной клетки. Проще представить себе, что на какой-то стадии развития капилляр располагается в непосредственной близости от поверхности тела нервной клетки, протоплазма которой при дальнейшем росте окружает капилляр со всех сторон н он, таким образом, оказывается включенным в нее.
Итак, мы видим, что в литературе существуют два взгляда на взаимоотношение нервных клеток и капилляров. Согласно первой точке зре-
ния, нервные клетки передвигаются по направлению к сосудам и группируются в непосредственной близости от источника питания. Иначе говоря, речь идет о периваскулярном расположении нервных клеток. Согласно второй точке зрения, тело нервной клешни рассматривается как центр, вокруг которого располагаются сосуды, т. е. речь идет о перицел-л юл я рном распределении капилляров.
Как же понять существование в центральной нервной системе того и другого способа взаимодействия нервных клеток и капилляров? С первого взгляда кажется вполне заданным считать нервные клетки как вы-сокодиференцированные элементы, функции которых находятся в теснейшей зависимости от непрерывного поступления к ним кислорода и питательных веществ, центрами притяжения других элементов. Это тем более вероятно, что сосуды представляют собой результат преобразования мезенхимных элементов, обладающих функцией передвижения. Но в то же время нельзя не считаться с тем, что в ряде случаев нервные клетки определенным образом стремятся к капиллярам.
Подвергнув этот вопрос анализу с точки зрения онтогенеза, мы нашли возможным сделать предположение, объясняющее различные способы взаимоотношений нервных клеток и капилляров. В 1947 г. нами была предложена, теория, рассматривающая развитие полушарий головного мозга в филогенезе и онтогенезе в зависимости от улучшения и смены систем, питающих мозг (кровеносной и ликворной). Представление о путях морфологического развития полушарий головного мозга обосновывалось сравнением результатов наших работ о развитии сосудистых сплетений мозга, исследований по кровообращению в мозгу и по развитию сосудисто-капиллярной сети в нем и т. д. Сопоставление данных этих работ с данными о развитии и миграции нервных клеток коры полушарий головного мозга и с результатами изучения развития глиоенюй ткани дало нам понять, каким образом в онтогенезе и филогенезе происходит смена ликворного питания нервных клеток ликворно-кровя-ным, а затем и кровяным.
Мы могли констатировать, что в процессе превращения поперечника мозгового пузыря в поперечник полушария головного, мозга взаимоотношения нервных клеток и капилляров изменяются несколько раз.
На одних этапах развития нервные клетки являются центрами притяжения капилляров, на других—клетки сами мигрируют к сосудам.
На ранней стадии развития передний мозговой пузырь состоит только из многослойного, матрикса и узкого, почти не содержащего клеток краевого покрова. Кровеносная система в этот период представлена широкими капиллярами, располагающимися в мягкой мозговой оболочке на наружной поверхности переднего мозгового пузыря (рис. 161). Мат-рикс в этом периоде развития сосудов не содержит. Па стадии существования бессосудистого матрикса клетки его получают питательные вещества из ликвора, выделяемого особо построенными сосудистыми сплетениями. Вслед за этим развитие поперечника переднего мозгового пузыря характеризуется тем, что клетки матрикса Многократно делятся, благодаря чему толщина всего слоя значительно увеличивается. Одновременно с этим отмечается врастание капиллярных сосудов в матрикс и образование в нем капиллярной сети. Таким образом, большое количество клеток, образующихся при делении клеток матрикса, нуждается в притоке большего количества кислорода и питательных веществ и в более быстром выведении продуктов обмена. Можно думать, что продукты обратного метаморфоза клеток матрикса в данный период являются стимулом, под влиянием которого совершается перемещение мезенхи-
мальных элементов в виде прорастающих капилляров по направлению к матриксу,
Во время прорастания капилляров в матрикс нервные элементы его некоторое время находятся в окружении тех веществ, которые поступают из жидкости, вырабатываемой сосудистыми сплетениями, и из крови, циркулирующей по капиллярной сети матрикса. По мере роста и превращения элементов матрикса происходит качественное изменение их обмена. Изменившиеся процессы обмена веществ требуют для нормального своего течения большого количества кислорода. Однако по сравнению с большим количеством клеток, содержащихся в матриксе, капил-
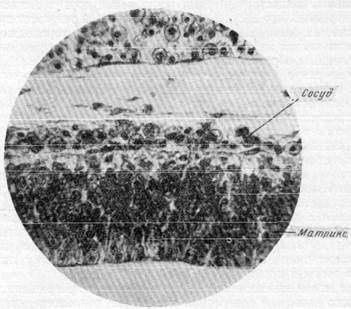
Рис. 161. Расположение кровеносных сосудов только в мягкой мозговой оболочке на ранке» стадии развития. Эмбрион крысы первой половины беременности (сагиттальный срез). Окраска по методу Массона. Увеличение 100.
ляров на данной стадии развития мало, следствием чего и является некоторое кислородное голодание, испытываемое клетками матрикса. Последнее обстоятельство приводит к тому, что нервные элементы матрикса начинают перемещаться, мигрируют по направлению к наружной поверхности мозгового пузыря, в мягкой мозговой оболочке которого располагается густая сеть артериальных сосудов. Как можно видеть на рис. 162, передвижение клеток матрикса происходит вдоль вертикально идущих отростков спонгиобластов (по направлению от полости мозгового пузыря к его наружной поверхности).
Миграция клеток приводит к образованию сначала так называемого кахалевсюго слоя, расположенного в непосредственной близости от наружной поверхности мозгового пузыря. Кислород и питательные вещества, содержащиеся в большом количестве в крови, циркулирующей по сети артериальных сосудов мягкой мозговой оболочки, на первых этапах создают наиболее благоприятные условия существования для переместившихся клеток.
Одновременно с этим известные изменения претерпевает также и кровь. Основное изменение заключается в превращении ядерных эритроцитов в безъядерные. Благодаря этому процессы обмена веществ в самом красном кровяном шарике крови становятся значительно менее выраженными и основная масса кислорода и питательных веществ переносится ими уже не для собственного потребления (Шейнис, 1949).
|
|
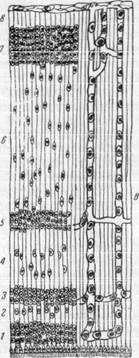
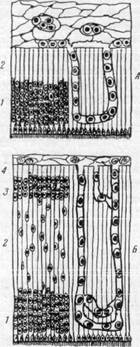
Рис. 162. Схема, иллюстрирующая образование коры и белого вещества полушария головного мозга.
Миграция нервных клеток и прорастание сосудов
в мозговое вещество.
А — стенка полушария головного мозга (эмбрион четырех недель): 1 — матрикс; 2 — краевой покров; Б — эмбрион середины третьего месяца: 1 — матрикс: 2 — промежуточный слой: 3 — закладка корковой пластинки; 4 — краевой покров. В — стенка полушария головного мозга на стадии 8 слоев (эмбрион 4—5 месяцев): 1 — матрикс; 2 — внутренний полосатый слой; 3 — внутренний переходный слой; 4 — наружный полосатый слой; 5 — наружный переходный слой; 6 — промежуточный слой; 7 — кора: 8 — краевой покров.
Все увеличивающаяся масса клеток, перемещающаяся к наружным частям стенки переднего мозгового пузыря, требует для своей жизнедеятельности все большего количества кислорода и питательных веществ. Вместе с тем передвижение клеток сопровождается изменением протекающих в них процессов обмена веществ, т. е. диференцировкой клеток матрикса и превращением их в различные элементы нервной ткани. Все это имеете взятое приводит к тому, что клеткам налипает нехватать того количества кислорода и питательных веществ, которые поступают к ним из артериальных сосудов мягкой мозговой оболочки. Недостаток кислорода находит отражение в гибели части мигрировавших клеток. Продукты обмена веществ оставшихся клеток и продукты распада по-
гибших клеток (главным образом их ядер) являются стимулом, под действием которого начинается бурное массовое врастание сосудов в стенку мозгового пузыря из мягкой мозговой оболочки, наблюдающееся нами во второй половине беременности.
Таким образом, на протяжении всего периода онтогенетического развития несколько раз происходит смена ориентации между нервными клетками и капиллярами. На одних этапах формирования эмбриона изменения, происходящие в процессах обмена веществ нервной клепки, выбывают рост капилляров по направлению к нервным клеткам, на других — изменившиеся и изменяющиеся нервные клеши сами перемещаются к источнику питания. С этой точки зрения понятно, почему в мозгу взрослого животного и человека можно встретить одновременно несколько нервных клеток в одной капиллярной петле и одну нервную клетку в отдельной капиллярной петле. При миграции нервных элементов целый ряд нервных клеток может оказаться расположенным в одной капиллярной петле. Дальнейший рост и диференцировка могут привести к тому, что одни клепки этой труппы станут более интенсивно функционировать, другие — менее. Повышение функции требует усиления процессов обмена веществ, что в свою очередь приводит к росту капилляров, направляющихся именно к той клетке, нагорая предъявляет наиболее высокие требования на кислород и питательные вещества. Таким образом, отдельная нервная клетка может оказаться лежащей в капиллярной петле или даже в нескольких капиллярных петлях. Приведенные факты показывают, что взаимное расположение нервных клеток и капилляров определяется телом нервной клетки.
Таким образом,подтверждается точки зрения И. П. Павлова о физиологической роли отдельных частей нервной клетки.
В одной из своих статей «Здоровое и больное состояние больших полушарий» (т. III, стр. 356, 1949) И. П. Павлов говорит: «Замыкание и образование новых связей мы относим за счет функций разделительной мембраны, если она существует, или просто утончающихся разветвлений между нейронами, между отдельными нервными клетками. Колебания возбудимости, переход, в тормозное состояние приурочиваем к самим нервным клеткам. Это размещение функций представляется нам вероятным в силу факта, что в то время мак новые связи, хорошо выработанные, очень долпо сохраняются, изменения возбудимости, переход в тормозное состояние суть очень подвижные явления. Явления возбуждения и торможения нам кажутся разными фазами в деятельности клеток коры больших полушарий».
Но существует и иная точка зрения на функциональное значение отдельных частей нервной клетки. Согласно этой точке зрения, основной центр нервной деятельности переносится с тела нервной клетки на синап-тический аппарат или на нейропиль, т. е. на густое сплетение тончайших разветвлений аксонов, переплетенных с многочисленными разветвлениями дендритов (Херрик и др.).
Имеются попытки со стороны ряда исследователей, изучающих кровоснабжение нервной системы, подтвердить эту теорию на основании степени снабжения капиллярами нейропиля и синапсов вообще в сравнении с телами нервных клеток. Эти авторы пытаются доказать, что нейропиль и синапсы снабжаются капиллярами интенсивнее, чем тела нервных клеток [Шаррер, 1937, 1944, 1945; Дунинг и Вольф (Dunning a. Wolf), 1937]. Так, например, Шаррер считает, что в тех случаях, когда синапсы располагаются вдали от тела клетки ,и в своей совокупности образуют нейропиль (первый слой коры больших полушарий), последний
и является местом сосредоточения наибольшего количества капилляров. Однако не каждый нейропиль обладает хорошей васкуляризацией. Так, например, нейропиль молекулярного слоя мозжечка имеет крайне незначительное количество капилляров. Для объяснения этого несоответствия в капиллярном снабжении нейропилей Шаррером было выдвинуто предположение о зависимости густоты капиллярной сети от количества митохондрий, содержащихся в синапсах. Специальные исследования, предпринятые в этом направлении, показали, что количество митохондрий различно в различных синапсах. Существуют нейропили, в синапсах которых число митохондрий крайне незначительно и вместе с тем есть нейропили с большим содержанием их. Поскольку по некоторым данным митохондрии являются местом сосредоточения дыхательных энзимов, Шаррер полагал, что синапсы с большим числом митохондрий в них могут быть отнесены к областям с наибольшим потреблением кислорода, а следовательно, и к участкам нервной системы — с наибольшей васку-ляризацией.
Исследователи,переносившие основную функциональную деятельность с тела нервной клетки на синапсы, видели подтверждение своей точки зрения в сравнении кровоснабженияузла тройничного нерва и коры. Кленки узла тройничного нерва посылают один из своих отростков на периферию, а другой в варолиев мост и продолговатый мозг и не имеют на своих телах синапсов. В то же время эти узлы содержат мало капилляров в отличие от мощной васкуляризации коры, имеющей огромное количество синапсов (Дунинг и Вольф, 1937).
Эта теория, переносящая центр основных процессов жизнедеятельности с тела нервной клетки в синапсы, может привлекать только своей новизной. Что же касается предпосылок, взятых для ее обоснования, то они в большинстве своем представляют собой пример логической непоследовательности и ложных фактов.
Основная масса данных, служащий для обоснования разбираемой теории, получена при изучении мозга костистых рыб, молодых аллигаторов и опоссумов. В мозгу указанных животных мы имеем или замкнутые желудочки мозга, где питание происходит за счет секрета сосудистых сплетений (рыбы и аллигаторы), или конечные артерии (опоссум).
Другими словами, условия, в которых осуществляется жизнедеятельность нервных клеток полушарий головного мозга, в этих случаях совершенно особые по сравнению с условиями у тех же клеток млеко-питающих с иным способом ликвообразования и кровообращения в мозгу.
Искусственность этой теории видна также и в том, что создается впечатление о постоянном сосредоточении синапсов на некотором расстоянии от тела клетки. На самом же деле во многих случаях синапти-ческие окончания располагаются на теле нервной клетки, так что трудно с этой точки зрения установить причину скопления капилляров вокруг клетки.
Нужно думать, что высокий индекс капилляров может зависеть и от тела самой нервной клетки, и от находящихся в ней синапсов. Что касается данных Дунинга и Вольфа, то они получены несовершенной методикой — инъекцией сосудов берлинблау с приготовлением тонких срезов и подсчетом капилляров на отдельных срезах. Кроме того, в их работе васкуляризация коры сравнивается с васкуляризацией узла тройничного нерва. При этом сравнивается кора, уровень метаболических процессов в которой чрезвычайно высок, с нервным узлом, функциональная деятельность клеток которого низка. Известно также, что клетки коры переносят кислородное голодание только в течение 5 минут, тогда
как клетки гассерова узла погибают после анемии, продолжающейся значительно больший промежуток времени.
При сравнительном изучении клеток узла тройничного нерва и клеток мезэнцефалического корешка тройничного нерва Е. Г. Балашева (1950) в нашей лаборатория показала, что оба названные образования хорошо снабжены кровью и имеют густую капиллярную сеть. Каждая нервная клетка узла тройничного нерва и мезэнцефалического корешка оплетена капиллярами. Тесное взаимоотношение тела нервной клетки мезэнцефалического корешка и капилляров выражается в том, что капилляры плотно прилежат к поверхности тела клетки. Подобное же соотношение между телом нервной клетки и капиллярами наблюдается в узле тройничного нерва, особенность кровоснабжения клеток которого заключается в том, что капилляры отделены от тела клетки, покрывающей ее соединительнотканной оболочкой.
Известно, что клетки обоих образований происходят из одного зачатка, имеют униполярную форму и характеризуются отсутствием синап-сов на их теле. Несмотря на отсутствие синапсов на теле, клетки того и другого образования хорошо снабжены кровью. В другом ряде опытов Е. Г. Балашева сравнивала кровоснабжение клеток узлов тройничного и блуждающего первое с кровоснабжением клеток симпатических узлов. Оказалось, что симпатические узлы, клетки которых имеют вокруг большое количество синапсов, очень скудно снабжены капиллярами. Так, в одной капиллярной петле лежит обыкновенно насколько нервных клеток с окружающими их синапсами. Таким образом, полученные в нашей лаборатории данные показывают всю неосновательность утверждения о зависимости количества капилляров от количества синаптических связей.
Изучение васкуляризации центральной и периферической нервной системы подтверждает, что главным центром метаболических процессов и функциональной деятельности или, как говорит И. П. Павлов, местом, где разыгрываются процессы возбуждения и торможения, является тело нервной клетки.
Подводя итог вышесказанному, мы считаем, что на настоящем уровне наших знаний нет никаких данных для того, чтобы переносить центр метаболических процессов нервной клетки из ее протоплазмы в парапластическую субстанцию, где расположены синапсы. На основании изучения резистентности нервных клеток к аноксемии мы можем оказать, что чем больше интенсивность процессов обмена веществ в той или иной группе нервных клеток, тем больше и кровоснабжение в области их расположения. Соответственно этому аноксемия прежде всего оказывается на тех ядерных группах ,и нервных клетках, которые обильно ва-скуляризованы.
Иначе говоря, степень кровоснабжения того или иного участка нервной ткани отражает напряженность функциональной деятельности нервных клеток, сосредоточенных в данной области центральной нервной системы.
Дата добавления: 2015-05-16; просмотров: 1399;