ВОЗДЕЙСТВИЯ 1 страница
Циркуляция крови в мозгу по сравнению с циркуляцией ее в других органах обладает рядом существенных особенностей.
Как известно, снабжение кровью большинства внутренних органов осуществляется по специальной артерии, проникающей через так называемые ворота органа в его паренхиму.
В своем дальнейшем ходе питающая артерия отдает по всем направлениям многочисленные ветви, в конце концов распадающиеся на капилляры. Таким образом, снабжение кровью в этих случаях происходит по направлению от центра к периферии.
Иные соотношения наблюдаются в мозгу.
Крупные артерии, представляющие собой ветви стволов, отходящих от виллизиева круга, располагаются на наружных поверхностях мозга. Как от крупных стволов поверхностных артерий, так и от их ветвей различного порядка вплоть до самых мелких под прямым углом отходят радиальные артерии, проникающие в мозговое вещество для снабжения кровью коры и белого вещества.
Ветви радиальных артерий в различных слоях коры и белого вещества распадаются на капилляры, благодаря чему каждый слой содержит не только более или менее крупные ветви той или другой радиальной артерии, но и густое капиллярное сплетение.
Таким образом, в отличие от других органов кровоснабжение мозга идет от периферии к центру.
Вторая особенность кровообращения в мозгу заключается в том. что в мозгу нельзя отметить тех определенных соотношений между питающей артерией и отводящими венами, которые обычно наблюдаются в других органах и особенно отчетливо в конечностях. Из изложенного в главе, посвященной анатомии артерий и вен головного мозга, следует, что на поверхности мозга артерии не только не сопровождаются венами, но часто идут совершенно в противоположных направлениях.
Так, например, вены, собирающие кровь с верхней наружной поверхности мозга, несут кровь к верхнему продольному синусу, т. е. идут по направлению от наружной к медиальной поверхности мозга. В то же время артерии, представляющие собой ветви передней мозговой артерии, направляются навстречу им от медиальной поверхности полушария на наружную поверхность его.
То же наблюдается и в отношении направления хода ветвей средней мозговой артерии и вен, отводящих кровь с наружной поверхности мозга в среднюю мозговую вену.
Еще более сложные взаимоотношения между артериями и венами наблюдаются в мозговом веществе.
|
Третьей особенностью мозгового кровообращения является характер реакций сосудисто-капиллярной сети мозга в ответ на ряд различных физиологических и патологических воздействий. Так, например, действие асфиксии на весь организм в целом сопровождается расширением сосудисто-капиллярной сети мозга и в то же время асфиксия вызывает сужение сосудов за малым исключением почти во всех органах тела.
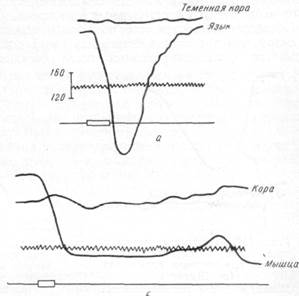
| Рис. 126. а — изменение скорости тока крови в сосудах теменной области коры и языка у кролика при раздражении головного конца перерезанного симпатического нерва (по Шмидту); б — изменение скорости тока крови в сосудах коры и мышцы при раздражении головного конца перерезанного симпатического нерва (по Шмидту). |
Особенности реакций сосудистой системы мозга выявляются и при воздействии на нее симпатической и парасимпатической системы. Раздражение симла-тического нерва на шее животного сопровождается сужением крупных по калибру артерий мягкой мозговой оболоч-ки всего лишь на 8— 10% от исходного диаметра. Но раздражение той же самой интенсивности вызывает сужение сосудов уха более чем на 50%.
На рис. 126, а, б показано, что раздражение головного конца перерезанного симпатического нерва имеет своим следствием резкое уменьшение скорости тока крови в сосудах языка и в mus. mylohyoideus кролика и в то же время вызывает лишь незначительные изменения скорости тока крови в сосудах теменной области.
Введение в ток крови симпатикотропных веществ, например, адреналина, также сопровождается различными реакциями сосудов мозга и периферии.
Как можно видеть из рис. 127, скорость тока крови в сосудах мозгового вещества увеличивается, тогда как в мышце она резко уменьшается. Противоположный эффект наблюдается в этом случае в условиях неизменяющегося общего кровяного давления и, следовательно, не зависит от него.
Различное влияние оказывает также перерезка симпатического нерва на шее животного. Освобождение от влияния со стороны симпатического отдела нервной системы имеет своим следствием резкое увеличение
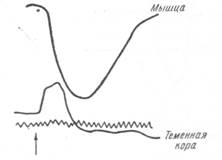
скорости тока крови в сосудах мышц. Интракраниальный ток крови при этом не изменяется совсем или меняется очень мало (рис. 128).
Одни и те же дозы веществ, расширяющих сосуды (нитроглицерин, кофеин и т. д.), вызывают значительно меньшее увеличение скорости тока крови в мозговых сосудах, чем в сосудах экстракраниальной области.
Наконец, особого внимания заслуживает чрезвычайная чувствительность нервных клеток мозговой ткани к недостатку кровоснабжения и связанная с этим тесная зависимость жизнедеятельности нервных элементов от непрерывного поступления к ним крови.
Последнее обстоятельство послужило основанием для предположения о том, что постоянная активность мозга должна быть связана с непрерывным функционированием всего сосудисто-капиллярного русла
|

|
Рис. 127. Изменение скорости тока крови в сосудах коры и мышцы при введении адреналина
(по Шмидту). Момент введения показан стрелкой.
Рис. 128. Изменение скорости тока крови в сосудах мозга и мышцы при перерезке симпатического нерва на шее животного (по Шмидту).
мозга в целом. Вследствие непрерывной потребности нервных клеток в определенном уровне напряжения кислорода в крови все капилляры мозга должны быть открыты. Крайним выражением данного взгляда явилось положение о наличии на капиллярах мозга эластической оболочки, не допускающей изменения их диаметра ни в сторону сужения, ни в сторону расширения (Е. К. Сепп, 1927).
В настоящее время последнее предположение отвергнуто всеми исследователями, так как накопленные факты не позволяют сомневаться в. том, что капилляры мозга могут изменять свой просвет в зависимости от многих физиологических и патологических воздействий. Установлено также, что гистологическое строение мозгового капилляра ничем особенным не отличается от строения его в других органах и тканях.
Специальные исследования, предпринятые в этом направлении П. Е. Снесаревым, позволили ему притти к заключению о наличии на стенке мозгового капилляра лишь разрозненных аргирофильных волокон, да и то появляющихся, начиная с определенного возраста. К тем же выводам пришел Д. С. Курбаналиев (1935), утверждающий, что в детском и молодом возрасте эластической оболочки на капиллярах мозга нет. Появление эластических волокон в стенке мозгового капилляра отмечалось этим автором только в старшем, а чаще в старческом возрасте и являлось с его точки зрения результатом изменений функционального характера.
Таким образом, стенка мозгового капилляра представляет собой чрезвычайно тонкую мембрану, состоящую из протоплазмы эндотелиаль-
ной клетки. Благодаря отсутствию оболочек на мозговом капилляре, нормальная, не измененная патологическим процессом стенка его подвижна и просвет капилляра мозга может изменяться в ту и другую сторону.
Как уже было сказано, малая устойчивость нервных клеток при условии недостаточного снабжения их кислородом указывает на особый характер функционирования всей сосудисто-капиллярной сети мозга в нормальных условиях.
Постановка вопросов о сущности циркуляции крови в мозгу имеет тем большее значение, что, как известно, те же вопросы для ряда других органов и тканей стоят совершенно в иной плоскости.
В отношении выяснения сущности циркуляции крови в органах и тканях работающих и находящихся в покос интересны опыты Крога (Krogh, 1929).
Крог инъицировал в кровяное русло при жизни лягушки или морской свинки китайскую тушь. В результате произведенных опытов он получил возможность составить достаточно точное сравнительное представление о количестве открытых и закрытых капилляров в различных органах к моменту смерти. При изучении препаратов, приготовленных из ряда органов, Крог установил различную степень заполнения сосудистого русла, что в свою очередь являлось косвенным доказательством различного количества открытых в момент инъекции капилляров.
В результате проведенных опытов было выяснено, что наибольшее количество открытых капилляров содержится в мозгу, печени и коже, тогда как в языке, а также в пустом желудке и кишках большинство капилляров закрыто. В соответствии с этим язык, пустой желудок и кишки инъицировались чрезвычайно мало, мозг, печень и кожа — всегда хорошо.
Различные мышцы тех же животных обнаружили различную степень инъекции. В одних покоящихся мышцах (например, в m. extensor tarsi у лягушки) количество заполненных тушью капилляров не превышало 5 на площади, равной 1 мм2. В других (например, m. rectus abdominalis) число открытых капилляров колебалось от 115 до 180 на той же площади и составляло 30—40% от общего количества капилляров, характерного для данных мышц.
Но совершенно иные соотношения наблюдались в тех случаях, когда в момент инъекции производилось раздражение того или иного органа или тетанизация мышц. Мало инъицирующийся в покойном состоянии язык лягушки значительно лучше инъицировался в условиях раздражения. Число инъицированных капилляров в m. extensor tarsi при раздражении мышцы увеличивалось с 5 до 195 на 1 мм2. Увеличение силы раздражения имело своим следствием раскрытие большего количества капилляров, благодаря чему наибольшая степень инъекции отмечалась после тетанизации мышц.
Если в покоящейся мышце лягушки среднее расстояние между открытыми капиллярами составляло 200—800 м, то после сокращения мышцы оно уменьшалось до 60—70 м. Различие в количестве открытых капилляров в работающих и покоящихся мышцах выступало отчетливо также при сравнении постоянно работающей мышцы диафрагмы с мышцей брюшной стенки, находящейся в покое. В то время как в первой можно было насчитать 2 500 открытых капилляров на площади в 1 мм2, на той же площади второй мышцы их было всего около 200.
Приведенные данные позволяют убедиться, что кровообращение ряда органов тела и мышечной системы находится в полном соответствии
с их функциональной деятельностью. Усиление функциональной деятельности влечет за собой увеличение числа открытых капилляров. Уменьшение функциональных требований, предъявляемых в каждый данный момент к тому или другому органу или мышце, влечет за собой полное закрытие части капиллярного русла.
Следовательно, в отношении некоторых органов и мышц не существует никакого сомнения в том, что капилляры в них могут становиться полностью непроходимыми для тока крови в течение длительного периода покойного состояния. При этом закрытие даже большого количества или большинства капилляров не сопровождается дегенерацией ткани органа или мышцы.
Другими словами, при таком способе кровообращения мы имеем дело со сменной циркуляцией. Сменность определяется в этих случаях не сужением и расширением капиллярного русла, а закрытием части его в покойном состояний и открытием при функционировании соответствующего органа или мышцы.
Что же представляет собой циркуляция крови в мозгу? Рассмотрим вначале кровоснабжение мозга тех животных, у которых оно осуществляется наиболее примитивным образом. Выше уже упоминался особый способ кровоснабжения мозга опоссума. Анатомическое распределение конечных артерий в мозговом веществе указанного животного создает такое расположение капилляров, при котором каждый обслуживает нервные клетки в радиусе не более 25 м от него (например, в коре мозжечка).
Отсутствие экспериментальных данных не дает возможности сделать, какое-либо заключение о реакциях капиллярной сети мозга опоссума при воздействии на нее физиологических или патологических факторов. Иначе говоря, мы не знаем, имеет ли место при указанных воздействиях расширение или сужение капилляров. Однако точное соответствие количества капилляров количеству и функциональным требованиям нервных клеток исключает всякое предположение о возможности полного закрытия капилляров с сохранением нормальных условий жизнедеятельности нервной ткани. Поэтому все капилляры мозга опоссума всегда открыты/ Каково же поведение капилляров мозга животных, для которых характерна непрерывная сосудисто-капиллярная сеть?
Исследование скорости тока крови в сосудах мозга в условиях закрытого черепа, а также наблюдения сосудов мягкой мозговой оболочки через «окно», герметически вставленное в череп, привело ряд исследователей к выводу о постоянстве циркуляции крови в мозгу (Clark a. Wenst-ler, 1938; Forbes a. Cobb, 1938; Lennox a. Gibbs, 1938). Исходя из того, что мозг всегда активен и его функциональные потребности близки к максимальным, эти исследователи считают, что все капилляры мозга всегда открыты.
При изучении циркуляции крови по сосудам мягкой мозговой оболочки с помощью капилляроскопа через «окно», герметически вставленное в череп, мы неоднократно имели возможность убедиться в том, что находившиеся под наблюдением сосуды различного диаметра никогда не изменяли своих размеров. Это в равной мере могло быть отмечено как в случаях наркотизации животных, так и в тех случаях, когда наблюдения производились на ненаркотизированных животных.
Таким образом, при отсутствии внешних раздражений, специфических для областей мозгового вещества, находящихся под наблюдением, сосуды мягкой мозговой оболочки этих областей спонтанно не расширя-
ются и не суживаются. И в этом отношении наши данные вполне совпадают с результатами работы Кларка (Clark a. Wenstler, 1938). Как известно, Кларк со своими сотрудниками изучал поведение сосудов мягкой мозговой оболочки теменной области у ненаркотизированного кролика через окно в черепе. Герметически вставленное «окно» обеспечивало нормальное существование сосудов мозга в закрытом черепе.
Как было установлено в этих опытах, продолжавшихся в течение нескольких месяцев, артерии и вены, располагавшиеся в мягкой оболочке, не изменяли в поле зрения свой просвет при умеренных колебаниях;
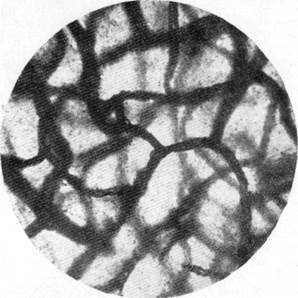
Рис. 129. Капиллярная сеть коры нормального животного, убитого декапиталией.
Импрегнация капиллярной сети мозга кошки по методу В. Н. Клосовского. Увеличение 400.
окружающей температуры, а также при шумах или звуках. Наряду с отсутствием спонтанных изменений просвета сосудов мягкой мозговой оболочки, сосуды уха обнаруживали колебания своего просвета в широких пределах даже при незначительных изменениях в окружающей среде.
Однако некоторые данные нашей лаборатории заставляют с осторожностью говорить о раскрытии в мягкой мозговой оболочке всех без исключения сосудов. Изучение артериальной сети мягкой мозговой оболочки в нормальных условиях и при асфиксии у животных без наркоза позволяет отметить значительно большее количество сосудов диаметром в 10—20 м в условиях асфиксии по сравнению с числом их у того же животного до опыта. В случае подтверждения полученных фактов можно будет говорить о закрытии некоторой части сосудов незначительного размера в мягкой мозговой оболочке в условиях нормального функционирования мозга при отсутствии внешних раздражений.
С аналогичными фактами сталкиваемся мы и при изучении капиллярной сети мозга нормального животного, убитого мгновенной декапн-гацией. Как можно видеть на рис. 129, капиллярная сеть мозга нормального животного, убитого декапитацией, характеризуется равно-
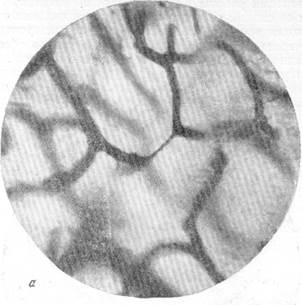

Рис. 130.Различные виды закрытых капилляров в капилярной сети коры нормального животного, убитого декапитацией.
а - закрытый капилляр в мозгу кошки; б — закрытый капилляр в мозгу собаки. импрегнация капиллярной сети по методу Б. Н. Клосовского. Увеличение 400.
мерностью просвета капилляров во всех отделах коры мозга; одинаковые по своему диаметру на всем своем протяжении капилляры формируют сеть, состоящую из правильных петель.
Обращает на себя внимание «округлость» колен капилляров, являющихся сторонами капиллярной петли. Такого рода форма капилляров говорит о напряжении капиллярной стенки, о хорошем тоническом состоянии ее в силу полного заполнения капилляра кровью и нормального давления в нем.
Изучение многих серий препаратов мозга нормальных животных, обработанных предложенным нами методом импрегнации, позволило убедиться в том, что все капилляры мозгового вещества в нормальных условиях открыты. Но среди правильных по форме и диаметру капилляров у нормальных животных можно встретить и то, что мы в общей форме называем закрытыми капиллярами. Закрытые капилляры в коре и белом веществе встречаются у нормального животного редко. На несколько полей зрения при всех поворотах микрометрического винта при увеличении в 400 раз можно отметить один и в исключительных случаях два закрытых капилляра.
Что же мы называем закрытым капилляром?
Выше уже говорилось о том, что у нормального животного, как правило, все капилляры имеют обычно один и тот же диаметр, различный в зависимости от вида животного. У кошки диаметр мозгового капилляра приблизительно равняется 6,8 м, у собаки он несколько больше 7 м, у человека равен приблизительно 8 м и т. д. Интересующие нас закрытые капилляры в своем поперечнике обычно не превышают 2 м.
Как можно убедиться из рис. 130, а, закрытый капилляр обычно представляет собой плотный тяж диаметром в 1—2 м, тянущийся от одного открытого капилляра к другому. Довольно часто встречаются закрытые капилляры, имеющие вид очень тонкой нити, как это показано на рис. 130, б. Длина описываемых образований весьма различна и в общем соответствует обычной протяженности мозгового капилляра.
Большей частью места отхождения закрытого капилляра от двух других, которые он соединяет, ничем не замечательны, но иногда в этих участках наблюдаются какие-то «наплывы» или скопления плазмы. Эти участки обычно имеют светлокоричневый цвет, тогда как открытые капилляры и соединяющий их закрытый капилляр интенсивно импрегниру-ются и представляются совершенно черными. Природа описанных образований не ясна и требует специального исследования. Можно думать, что это остатки протоплазмы закрывшегося капилляра, но можно также предполагать, что в этих местах располагаются эндотелиальное, а иногда и адвентициальное ядра капилляра.
Приведенный здесь фактический материал позволяет поставить в порядке обсуждения вопросы о том, что же представляют собой описанные образования: 1. Соединительнотканную связку или эндотелиальный мостик между капиллярами? 2. Временно закрытый, не функционировавший в момент смерти животного, но способный функционировать капилляр? 3. Капилляр, уже неспособный раскрыться, т. е. капилляр атрофирующийся?
Оставляя два последних вопроса для рассмотрения их в последующих работах, мы остановимся здесь только на первом.
В работах отдельных исследователей нам удалось отыскать описание особых образований, встречающихся в капиллярной сети центральной нервной системы, несколько напоминающих рассматриваемые здесь закрытые капилляры. Так, например, Ранке (1915) сообщает о наблю-
давшихся им в различных частях центральной нервной системы плотных, содержащих фибриллы «соединительных мостиках». Эти «соединительные мостики», состоящие, по мнению Ранке, из адвентициальной плазмы, большей частью встречались в капиллярной сети серого вещества спинного мозга и лишь в редких случаях наблюдались в коре полушарий.
Придавая большое значение этим образованиям, Ранке отводил им значительную роль при патологических процессах в мозгу. Так, например, он считал, что фибробластическая организация при размягчениях мозга у человека может происходить и без участия зернистых шаров лишь на основе «соединительных мостиков». За счет последних он относил также образование фибробластических адвентициальных сетей, подобных тем, которые описаны П. Е. Снесаревым при параличе, артериосклерозе и т. д.
Плотные мостики, располагающиеся между капиллярами, еще до Ранке были описаны Генле. Наблюдал их также и Рамон Кахал, назвавший их «cordons unitifs». К сожалению, в доступной нам работе Ранке нет иллюстраций. Поэтому трудно судить, о каких образованиях идет здесь речь и являются ли они аналогичными тем, которые мы считаем закрытыми капиллярами.
В книге Е. К. Сеяна «Клинический анализ нервных болезней», ч. Г (1927) приведен рисунок Мингацинни, на котором изображена капиллярная сеть участка мозга прогрессивного паралитика. На этом рисунке видны плотные, не имеющие просвета тяжи, соединяющие соседние капилляры. По своему внешнему виду тяжи напоминают то, что мы называем закрытыми капиллярами. По мнению Мингацинни, наличие такого рода плотных тяжей в капиллярной сети мозга должно говорить о протекающих здесь процессах обратного развития. К этому же мнению присоединяется и Е. К. Сепп, считающий плотные тяжи специальными приспособлениями, возникающими в капиллярной сети мозга в случаях затрудненного кровообращения в мозгу.
Таким образом, ряд исследователей наблюдал и описал под различными названиями плотные тяжи в капиллярной сети мозга неизвестной природы. Многие связывали их появление с процессами обратного развития, протекающими в организме при патологических состояниях, и считали их соединительнотканными мостиками, укрепляющими отдельные части капиллярной сети.
Для того чтобы решить вопрос о характере плотных тяжей, описанных предшествовавшими авторами в патологически измененной капиллярной сети мозга человека и обнаруженных нами также и в капиллярной сети нормальных животных, мы предприняли опыты с прижизненной инъекцией.
Введение трипановой сини в ток крови нормального животного при жизни его дало нам возможность убедиться, что среди равных по диаметру мозговых капилляров располагаются отдельные резко суженные капилляры, имеющие вид изображенного на рис. 130, а. Диаметр этих капилляров в 3—4 раза меньше диаметра нормально открытых капилляров. Наиболее характерным является тот факт, что резко суженные капилляры содержат окрашенную плазму. Это обстоятельство указывает на имеющийся в суженных капиллярах просвет. Другими словами, мы имеем дело с действительно капиллярами, резко сократившимися и проходимыми для плазмы, а не с соединительнотканными укрепляющими мостиками, как это предполагалось некоторыми исследователями. Установив природу этих образований, мы не можем, однако, указать на причину появления резко суженных капилляров в капиллярной сети нор-
мального животного. Факты, полученные нами при прижизненной инъекции трипановой сини в ток крови, указывают, что суженные капилляры в момент смерти нормального животного уже были выключены из общей капиллярной сети мозга. Резкое сокращение, вследствие которого они становились непроходимыми для красных кровяных телец, позволяет считать их нефункционирующими.
Нельзя пока сказать, временно ли закрываются капилляры или отдельные из них, подобные капилляру, представленному на рис. 130, б, в последующем совсем не открываются и должны быть отнесены к категории атрофирующихся. Можно лишь предполагать, что капилляры, имеющие на импрегнированных препаратах вид тончайшей нити (рис. 130, б), должны быть отнесены к числу капилляров с полностью закрывшимся просветом.
Таким образом, изучение капиллярной сети мозга нормальных животных, убитых мгновенной декапитацией, позволяет констатировать, что отдельные капилляры этой сети могут резко суживаться или даже закрываться в условиях отсутствия внешних раздражений.
Каковы же соотношения в сосудистой сети мягкой мозговой оболочки и капиллярной сети мозгового вещества нормальных животных при раздражении различных рецепторов, другими словами, — каково поведение сосудов в различных полях мозга в условиях различных функциональных требований, предъявляемых к расположенным в этих полях нервным клеткам.
В опытах с раздражением вестибулярного аппарата кролика введением в его ухо теплой или холодной воды или при вращении животного мы неизменно отмечали расширение сосудов мягкой мозговой оболочки в лобно-теменной области (Б. Н. Клосовский, 1942). Расширение было весьма значительно и составляло около 50% первоначального размера сосудов, измеренного с помощью окуляр-микрометра до опыта. Характерно, что увеличение диаметра сосудов, наблюдавшихся при помощи ка-пилляроскопа через «окно», вставленное герметически в череп, отмечалось только в указанной области. Через второе «окно», располагавшееся под мягкой мозговой оболочкой в затылочной области, можно было отметить некоторое сужение сосудов.
Следовательно, раздражение рецептора имело своим следствием расширение сосудов в участке мягкой мозговой оболочки, располагавшемся над областью мозгового вещества, нервные клетки которой обнаруживали повышенную жизнедеятельность.
Термопарой, погруженной в эту область, можно было зарегистрировать повышение температуры на 0,1—0,2° по сравнению с исходной. Это в свою очередь указывало на расширение сосудов внутримозгового вещества.
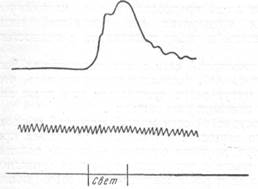
Аналогичный опыт, но с раздражением зрительного рецептора, был проделан Шмидтом (1936). Как видно на рис. 131, освещение глаза ку-рарезованной кошки в условиях искусственного дыхания имеет своим следствием увеличение скорости тока крови в сосудах затылочной области. Определяемое термопарой увеличение тока крови быстро возвращается к исходному уровню, как только закрывается глаз и прекращается раздражение.
В литературе имеется описание случая ограниченной angioma arte-riale racemosum, располагавшейся в левой затылочной доле. Аускульта-тивно в этом месте можно было услышать шум, который слышал и сам больной. Шум был значительно ослаблен, если больной находился в покойном состоянии, например, отдыхал с закрытыми глазами. Но как
.259
только больной начинал читать или пытался осмыслить виденное, шум резко усиливался. И в то же время другие виды мозговой деятельности, вызванные раздражением обонятельного, слухового или другого рецептора, увеличения шума не давали.
|
Приведенные эксперименты позволяют понять значение адэкватно-сти раздражителя при изучении изменения просвета сосудов в той или другой области мозга или мягкой мозговой оболочки. И с этой точки зрения могут вызвать возражения данные Кларка и Венстлера, установивших, как мы видели, неизменность просвета сосудов в мягкой мозговой оболочке теменной области у кролика при температурной и другой стимуляции.
| Рис. 131. Увеличение скорости тока крови в сосудах затылочной области при освещении глаза кошки. |
Это возражение заключается в том, что кролик является животным в основном оль-фактивным и тригеминальным. Поэтому обонятельные и вкусовые раздражения у него представляют собой значительно более сильные стимуляторы, чем слуховые или температурные. Кроме того, в опытах Л. А. Новиковой и Г. Я. Хволеса (1946) было установлено, что любое раздражение сопровождается у кролика увеличением электрической активности прежде всего на обонятельном тракте. Другими словами, каждое раздражение вызывает у кролика усиленную деятельность прежде всего отдела мозга, связанного с обонянием.
Отсюда ясно, что раздражение, применявшееся Кларком, не являлось адэкватным для той области мозгового вещества, сосудистая сеть мягкой мозговой оболочки которой изучалась через окно в черепе. При отсутствии же особых функциональных требований к той или другой области мозгового вещества трудно ожидать изменений просвета сосудов в соответствующем отделе мягкой мозговой оболочки.
Таким образом, результаты эксперимента Кларка и Венстлера вследствие методической ошибки не могут отрицать возможности расширения или сужения сосудов в областях мозга с повышенной в данный момент функцией.
Но эти эксперименты, как уже говорилось, совершенно правильно указывают на отсутствие изменений просвета сосудов в областях, не функционирующих активно.
Напротив, из результатов описанных опытов следует, что повышение функциональной деятельности в той или другой области мозга при раздражении специфического для этой области рецептора сопровождается увеличением кровообращения в соответствующей области мозга. Увеличение васкуляризации находит свое отражение в расширении капиллярного русла усиленно функционирующего участка мозгового вещества. Тут же возникает вопрос: как ведет себя капиллярная сеть мозга в участках, функциональная деятельность нервных клеток которых в данный момент понижена. Выше уже говорилось, что в наших опытах было отмечено сужение сосудов мягкой мозговой оболочки затылочной доли при одновременном расширении их в лобной доле.
Не ставя для себя целью исследование поведения сосудов в различных областях мозга при раздражении рецептора, специфического лишь для одной определенной области, мы ограничились в свое время лишь констатацией факта сужения сосудов. К сожалению, по этому вопросу в литературе не существует никаких экспериментальных данных. Можно найти лишь теоретические соображения отдельных исследователей (например, де Но), считавших возможным не только сужение сосудов в участках мозгового вещества с пониженной функциональной деятельностью нервных клеток, но и полное закрытие части капиллярного русла в них.
Дата добавления: 2015-05-16; просмотров: 917;