СНАБЖЕНИЕ НЕРВАМИ СОСУДОВ МЯГКОЙ МОЗГОВОЙ ОБОЛОЧКИ МОЗГА
Нервные волокна, идущие к сосудам мягкой мозговой оболочки и мозга, берут свое начало в двух источниках.
Первым является симпатическое сплетение внутренней сонной и позвоночной артерии.
Видимое невооруженным глазом сплетение нервных волокон, располагающееся на внутренней сонной артерии, было установлено еще в первой половине XVIII века. Однако единственный известный исследователям того времени метод анатомической препаровки давал возможность получить лишь общие представления о ходе и положении нервных стволов и их крупных подразделений в обнаруженном ими сплетении.
Работы последующего столетия, выполнявшиеся тем же способом, но усовершенствованным уже предварительной обработкой препаратов различными веществами и главным образом кислотами (уксусной, азотной), позволяли установить детально как расположение отдельных нервных стволов в сплетении, так и связь его с симпатическим нервным стволом.
Макроскопическое изучение нервов на внутренней сонной артерии и ее ветвях было дополнено затем микроскопическим исследованием тех же нервов, отпрепарованных после предварительной протравы с последующим окрашиванием (Белоусов, В. П. Воробьев, Кондратьев, Михайлов, Соколов и др.).
Тем не менее значительное усовершенствование методов не привело к накоплению достаточного количества фактов, на основании которых можно было бы сказать, что вопрос об иннервации сосудов мозга даже в той части, которая касается нервных сплетений на крупных артериях, снабжающих его кровью, решен полностью.
В литературе нет указаний на существование точных и прочно уста-новленных фактов, объясняющих происхождение того большого количества нервных волокон, которое можно наблюдать на сонных и позвоночных артериях. Еще в меньшей степени выяснено взаимодействие нервных стволов и волокон, входящих в состав сплетения с сосудистой стенкой, на которой они располагаются. До самого последнего времени не существовало подробного описания даже чисто анатомического распределения нервных стволов в сплетениях, располагающихся на артериях, подводящих кровь к мозгу, а также на сосудах основания головного мозга и их главных ветвях.
Опубликованная в 1948 г. работа А. С. Цветкова дает уже более детальное описание формы сплетений на внутричерепных отделах сонной и позвоночной артерии, а также на их основных ветвях вплоть до деления последних в мягкой мозговой оболочке.
Эта работа представляет собой наиболее полное и детальное описание сплетений по сравнению с предыдущими работами. В дальнейшем при рассмотрении морфологии и анатомии сплетений мы будет поэтому основываться на данных, сообщенных А. С. Цветковым.

|
Преганглионарные во
локна симпатической при
роды для сосудов головы
и шеи отходят от клеток
бокового рога спинного
мозга на уровне С8 и T1-3
(рис. 120), направляются
в звездчатый узел и верх
ний шейный симпатиче
ский узел, на клетках ко
торых они частично окан
чиваются. Таким образом,
крупный нервный ствол,
отходящий от верхнего
полюса верхнего шейного
симпатического узла,
| Рис. 120. Схема симпатической и парасимпатической иннервации сосудов мозга. |
включает в свой состав часть постганглионарных и часть преганглионарных волокон. Нервный ствол п. caroticus internus идет вместе с внутренней сонной артерией по каналу внутренней сонной артерии. Вместе с названной артерией п. caroticus internus входит в полость черепа, где ветвится на 1—3 ветви, которые являются основными стволами образующегося сплетения внутренней сонной артерии.
Эти 1—3 нервных ствола тянутся вдоль внутричерепного отдела внутренней сонной артерии, располагаясь в большинстве случаев на внутренней и наружной поверхности ее, и объединяются прямолинейными или дугообразными анастомозами, формируя сплетение.
При наличии в сплетении двух нервных стволов, из которых один идет по внутренней, а другой по наружной поверхности внутренней сонной артерии, отмечается большое количество вариаций в способе иннервации ветвей внутренней сонной артерии, систематизировать которые трудно.
В тех случаях, когда в образовании сплетения принимают участие три ствола (внутренний, наружный и задний), наблюдается преимущественное снабжение каждым из них определенной ветви внутренней сонной артерии.
Так, от внутреннего ствола нервные волокна отходят главным образом к передней мозговой и передней соединительной артерии, образуя сплетения, которые носят одноименные названия. Значительно меньшее количество волокон внутренний ствол дает для средней мозговой и задней
соединительной артерий.
|
От наружного ствола нервные волокна отходят преимущественно к средней мозговой артерии и лишь небольшая часть волокон этого ствола вступает в сплетение передней мозговой и задней соединительной артерии. Сплетение последней артерии в основном формируется за счет нервных волокон заднего ствола.
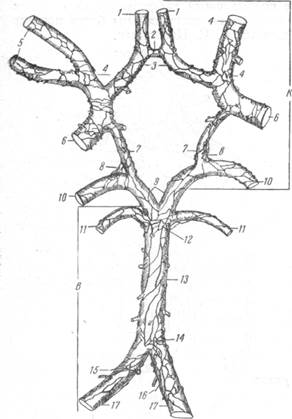
| Рис. 121. Распределение нервов «каротидной» и вертебральной системы» по артериям основания головного мозга (по Цветкову). К — «каротидная система» сосудистых нервов: 1 — артерии мозолистого тела; 2 — передняя соединительная артерия; 3 — передняя мозговая артерия; 4 — среднемозговая артерия; 5 — ветви среднемоз-говой артерии; 6 — внутренняя сонная артерия (внутричерепная часть); 7 — задняя соединительная артерия; 8 — переход сплетения задней соединительной артерии на каротидную часть задней мозговой артерии; 10 — задняя мозговая артерия (каро- тидная часть). В — «вертебральная система» сосудистых нервов: 9 — задняя мозговая артерия (базилярная часть); 11—верхняя мозжечковая артерия; 12 — переход сплетения основной артерии на верхнюю мозжечковую артерию; 13 — основная артерия; 14, 15, 16 — подходящие нервные стволы; 17 — позвоночные артерии. Нервы артерий основания мозга зарисованы со стороны, прилегающей к мозгу. |
Существует большое разнообразие в отношении протяжения обособленного хода того или другого крупного нервного ствола, входящего в состав сплетения какой-либо из артерий. В одних случаях нервный ствол может быть прослежен как целое не только на протяжении его хода вдоль внутренней сонной артерии, но и при переходе на одну из ее ветвей. В других - обособленность хода нервного ствола сохраняется только до места деления сонной артерии на среднюю, переднюю и заднюю соединительную артерии. Наконец, часто нервный ствол в месте деления артерии в свою очередь распадается на ветви, каждая из которых служит для образования сплетения той или другой из названных выше ветвей внутренней сонной артерии.
Сплетение задней соединительной артерии представляет собой связующее звено между нервными сплетениями внутренней сонной и позвоночной артерии.
Задняя соединительная артерия объединяет, как было сказано, два источника кровоснабжения головного мозга. Хорошо выраженное нерв-
ное сплетение этой артерии сформировано из нервных стволиков, достигающих 200 м в диаметре. Густое сплетение состоит из узких петель, вытянутых по длине сосуда. От сплетения отходит большое количество нервных веточек к задней мозговой артерии, что отчетливо представлено на приводимом здесь рисунке, взятом из работы А.С.Цветова (рис. 121).
|
Сплетение позвоночной артерии, имеющее источником своего происхождения звездчатый узел и шейные узлы симпатической нервной системы, включая в себя от 1 до 4 нервных стволов, располагающихся на передней и задней поверхностях сосуда. Как и нервы, входящие в состав сплетения внутренней сонной артерии, нервные стволы сплетения позвоночной артерии сохраняют свой обособленный ход на различном протяжении.
В ряде случаев можно видеть, как крупные нервные стволы доходят до места отхождения верхних мозжечковых артерий, но чаще обособленный ход того или иного крупного ствола сохраняется до места слияния позвоночных артерий в основную или даже на некотором протяжении вдоль последней. Вместе с тем иногда основные нервные стволы, входящие в состав сплетения позвоночной артерии, распадаются на ветви уже при входе этой артерии в полость черепа.
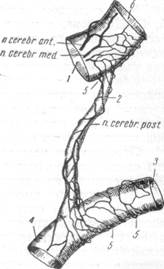
| Рис. 122. Переход сплетения задней соединительной артерии на каротидную часть задней мозговой артерии (по Цвет-кову). 1 — внутренняя сонная артерия, 2 — задняя соединительная арте рия; 3 — каротидная часть зад ней мозговой артерии; 4 — ба- зилярная часть задней мозговой артерии; 5 — ветви сплетения внутренней сонной артерии, при нимающие участие в образова нии сплетений задней соедини; тельной артерии и каротидной части задней мозговой артерии. |
Сплетение нервных волокон основной артерии представляет собой продолжение нервных сплетений слившихся позвоночных артерий. У места отхождения верхних мозжечковых артерий сплетение основной артерии меняет свой прямолинейный или спиральный ход на поперечный и, разделяясь, переходит в сплетение правой и левой верхних мозжечковых артерий. Лишь весьма незначительная часть тонких нервных ветвей отходит от сплетения основной артерии к базилярной части задней мозговой артерии (рис. 122).
Как видно на рис. 121 и 122, каротид-ный и базилярный участки задней мозговой артерии получают различное количество нервных волокон. При исследовании взаи-
моотношений сплетений внутренней сонной и позвоночной артерии рядом авторов отмечалось, что у человека сплетение задней мозговой артерии в основном является продолжением сплетения внутренней сонной артерии. Последнее обстоятельство, по мнению некоторых исследователей, должно указывать на вероятность иннервации сосудов всего большого мозга за счет нервов, отходящих от симпатического сплетения внутренней сонной артерии (Вильяме, 1936)1.
При сравнении густоты сплетения и числа нервных волокон с диаметром задней мозговой артерии обращает на себя внимание несоответ-
1 Иные соотношения отмечены у обезьян, у которых сплетение задней мозговой артерии формируется нервными волокнами периваскулярного сплетения основной артерий и, следовательно, сосуды, получающие кровоснабжение по задней мозговой артерии, иннервируются сплетением основной артерии.
ствие количества нервных ветвей, образующих сплетение, с просветом этого сосуда. В то время как задняя соединительная артерия, имеющая диаметр, наименьший по сравнению с другими артериями основания мозга, обладает густым сплетением нервных волокон, задняя мозговая артерия, с просветом значительно большим, получает лишь незначительное нервное снабжение.
Подобные соотношения объясняются тем, что нервное сплетение задней соединительной артерии является не только собственным сплетением этого сосуда, но и представляет собой транзитный путь для нервов, формирующих сплетение задней мозговой артерии.
Мы отсылаем интересующихся к уже упоминавшейся работе А. С. Цветкова, в которой представлены полные данные относительно направления хода, количества нервных стволов в сплетениях, расположения их по отношению к стенке сосудов, характера анастомозов, наличия узлов в сплетениях и морфологической формы последних.
Как следует из приведенных описаний, густые сплетения нервных волокон располагаются на внутренних сонных и позвоночных артериях, а также на их ветвях внутри черепа. Возникает вопрос: каковы же взаимоотношения массы нервных волокон со стенкой сосуда, на которой они распределяются?
Проведенные исследования не оставляют повода для сомнения в том, что часть волокон, идущих в составе сплетений вдоль сосудов, оканчивается в адвентиции их типичными концевыми чувствительными препаратами.
В литературе имеются сообщения, согласно которым мышечная оболочка сосудов получает чрезвычайно тонкие, нежные нервные волоконца, оканчивающиеся в ней. Большинство авторов в своих работах отмечает трудности прослеживания нервных волокон в мышечной оболочке (tunica muscularis). Эти авторы склонны рассматривать отдельные нервные волокна, наблюдающиеся в редких случаях, как результат косо прошедшего среза, вследствие чего нервные волокна, принадлежащие в действительности адвентиции, кажутся расположенными в средней оболочке сосуда (Н. Д. Довгялло, 1932).
Второй источник, служащий для образования нервных сплетений на сосудах основания головного мозга, впервые был обнаружен Белоусовым в 1899 г. Изучая нервы на сосудах, инъицированных затвердевающей массой, этот автор отметил ветвь, отходящую от глазодвигательного нерва. На протяжении приблизительно сантиметра эта ветвь шла вместе с задней мозговой артерией, затем распадалась на ветви меньшего диаметра и формировала сплетение с многочисленными веточками, отходящими к расположенным поблизости сосудам мягкой мозговой оболочки. Данные, полученные Белоусовым, впоследствии были подтверждены рядом авторов, в том числе П. Е. Снесаревым (1929) и др.
На большое постоянство участия ветвей глазодвигательного нерва в формировании сплетений на сосудах основания мозга в свое время указывал также Н. Д. Довгялло (1932). Он отметил тонкие нервные стволики, проходившие в направлении не только задней мозговой артерии, но и вступавшие в состав сплетений верхней мозжечковой артерии, артерии сосудистого сплетения, а также в небольшой своей части в сплетение основной артерии. Таким образом, давно установлен и многократно подтвержден факт отхождения от глазодвигательного нерва очень постоянных ветвей к мягкой мозговой оболочке и расположенным в ней сосудам. Однако, несмотря на общепризнанность этого положения, никем из исследователей до настоящего времени не установлена природа такого рода нервных волокон. Отсутствие гистологических
методов, пригодных для отличия миэлинизированных волокон парасимпатической природы от аналогичных волокон симпатического происхождения, не дает возможности отнести ту или другую группу их к симпатической или парасимпатической системе.
Для того чтобы установить деление нервных волокон на сосудах мягкой мозговой оболочки на симпатические и парасимпатические, мы решили проследить миэлинизацию сосудистых нервов на ранних этапах онтогенетического развития.
В центральной нервной системе и в периферических нервах нервные волокна покрываются миэлином не одновременно, а в зависимости от функции и филогенетической давности той или иной системы. Поэтому можно было ожидать, что не все нервы на сосудах мозга будут приобретать миэлиновую оболочку одновременно. Изучение миэлинизации нервных волокон на сосудах мозга подтвердило правильность нашего предположения, положенного в основу нашего исследования.
При рассмотрении серий непрерывных срезов, приготовленных методом Кульчицкого-Вольтерса (наиболее тонко выявляющим все миэлини-зированные волокна) из мозгов кошек различного возраста, было установлено, что первые миэлинизированные волокна на сосудах мозга появляются у котенка на 5—б-й день после рождения. На 9-й день эти :волокна уже могут быть отчетливо прослежены, начиная от места их возникновения от глазодвигательного нерва и далее на некотором протяжении хода в адвентиции сосуда, где они в конце концов и теряются. Приводимые здесь микрофотографии из мозга девятидневного котенка позволяют проследить это с большой наглядностью.
На рис. 123, а ясно видно, как от глазодвигательного нерва отходит пучок миэлинизированных волокон. Этот пучок нервных волокон еще с большей отчетливостью выступает на следующем срезе, представленном на рис. 123, б, на котором виден переход волокон от глазодвигательного нерва в адвентицию сосуда. Рис. 123, в позволяет проследить ход пучка миэлинизированных волокон в адвентиции сосуда. Не менее очевидно расположение волокон по ходу сосуда на рис. 123, г.
Характерно, что волокна, отходящие от глазодвигательного нерва к сосудам мозга, проходят стадии миэлинизации несколько позднее, чем другие волокна того же глазодвигательного нерва, что с очевидностью говорит об особой их функции.
Волокна пучка, отходящего к сосудам мозга от глазодвигательного нерва, могут быть прослежены лишь на небольшом протяжении их хода в адвентиции сосуда. Отмеченное обстоятельство служит указанием на то, что нервные волокна этого пучка оканчиваются на многочисленных нервных клетках, разбросанных в адвентиции сосудов, т. е. являются преганглионарными. Постганглионарные волокна того же сосудодвига-тельного пути являются аксонами клеток, в большом количестве обнаруживаемых в адвентиции по ходу сосудов, аксонами тех клеток, на которых оканчиваются преганглионарные волокна, идущие в составе глазодвигательного нерва.
Одним из интереснейших моментов является отсутствие на сосудах мозга 5—9-дневного котенка миэлинизированных нервных волокон, происходящих от других черепномозговых нервов или от симпатического нерва.
Миэлинизация вегетативных волокон парасимпатической природы всех других (за исключением глазодвигательного нерва) черепномозговых нервов для сосудов мозга, в том числе и для сосудов продолговатого мозга, начинается значительно позже. Следовательно, миэлинизированные
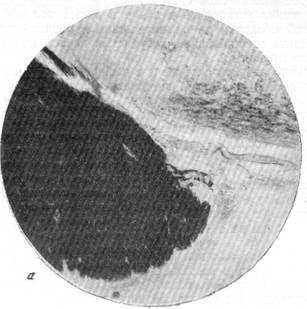

Рис. 123. Отхождение преганглионарных миэлинизированных волокон парасимпатической природы от корешка глазодвигательного нерва.
я — отхождение пучка парасимпатических волокон от корешка III нерва: б - те же волокна в момент перехода их в адвентицию сосуда
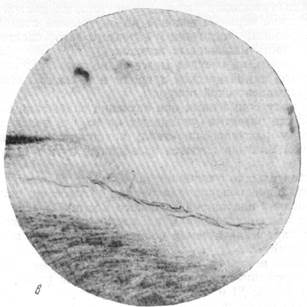
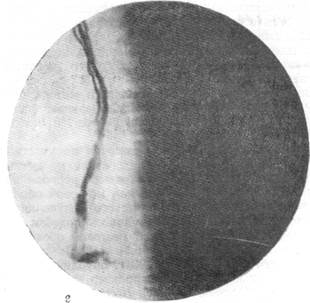
Рис. 123. Отхождение преганглйонарных миэлинизированных волокон парасимпатической природы от корешка глазодвигательного нерва.
в — ход тех же волокон в адвентиции сосуда при малом увеличении; г — ход тех же
волокон в адвентиции сосуда при большом увеличении.
Микрофотографии сделаны с 4 препаратов одной и той же непрерывной серии срезов.
Окраска по Кульчицкому.
волокна, обнаруженные нами на сосудах мозга 9-дневного котенка, являются нервными волокнами парасимпатической природы, идущими в составе глазодвигательного нерва. Кроме того, на данной стадии развития симпатические нервные волокна на внутренней сонной артерии еще не миэлинизированы, тогда как у взрослого это сплетение содержит значительную часть миэлиновых волокон.
Таким образом, на сосудах мозга 9-дневного котенка отсутствуют нервные волокна симпатической природы и имеются миэлинизированные волокна парасимпатической природы.
Данные, полученные нами при изучении снабжения нервами сосудов мозга в процессе развития, указывают на то, что волокна парасимпатической природы (сосудорасширители) приобретают миэлиновые оболочки, прежде чем миэлинизируются проводящие пути в полушариях головного мозга.
Это обстоятельство указывает на большое значение, которое имеют сосудорасширяющие волокна для регуляции просвета мозговых сосудов при формировании полушарий головного мозга.
Имеются также указания и на другие черепномозговые нервы как на источник парасимпатических волокон для сосудов мягкой мозговой оболочки и мозга.
Так, помимо ветвей от III пары черепномозговых нервов, отходящих к артериям основания головного мозга, описаны ветви, отходящие от VI, VII, VIII, IX, X, XI и XII пар черепномозговых нервов. Все эти нервные ветви принимают участие в образовании нервных сплетений на сосудах.
По некоторым указаниям, имеющимся в литературе, и V черепно-мозговой нерв является источником двигательных и чувствительных волокон для передней мозговой артерии. Так, Ларселл (Larsell, 1918) описал мякотные и безмякотные волокна, отходящие от тройничного нерва и заканчивающиеся в стенке передней мозговой артерии чувствительными и двигательными концевыми аппаратами. Эти данные, полученные на различных млекопитающих, не были, однако, подтверждены наблюдениями на человеке, на сосудах мозга которого нервных волокон от тройничного нерва отметить не удалось.
Естественно, что небольшие возможности метода препаровки ограничили исследования изучением сплетений нервных волокон лишь на основных артериях, снабжающих головной мозг, и отходящих от них крупных ветвях на основании мозга.
Прослеживая сплетения нервных волокон на участках этих ветвей, прилежащих к виллизиеву кругу, исследователи не могли получить каких-либо данных относительно расположения нервов на дальнейших разветвлениях мозговых артерий в мягкой мозговой оболочке.
Лишь при сочетании микроскопического изучения препаратов с окраской нервов метиленовой синью удалось подойти вплотную к рассмотрению иннервации сосудов мягкой мозговой оболочки.
Использовав предложенный Эрлихом (1885) метод окрашивания нервов на сосудах, Иванов уже в 1893 г. опубликовал работу, впервые ставившую своей целью проследить характер нервных окончаний в соединительной ткани оболочек. Касаясь расположения нервов на сосудах мягкой мозговой оболочки головного мозга, он отметил наличие большого количества мякотных и безмякотных нервных волокон, проходящих по обеим сторонам вдоль сосуда в виде толстых и тонких нервных стволиков. Такого рода нервное сопровождение сосудов, тем более выраженное, чем больше диаметр сопровождаемого сосуда, было прослежено Ивано-
вым вплоть до самых мелких или, как он говорит, «терминальных» разветвлений сосудов мягкой мозговой оболочки. Описанные безмякотные и более крупного диаметра миэлиновые волокна, как он мог наблюдать, образуют сплетения на поверхности сосудов. Иванов показал, что от сплетения на поверхности сосуда мягкой мозговой оболочки отходят нервные веточки, формирующие более глубокие сплетения, располагающиеся между адвентицией и мышечной оболочкой сосуда.
Таким образом, на самом раннем этапе развития учения об иннервации сосудов мягкой мозговой оболочки и мозга было уже описано большое количество нервных волокон на сосудах, расположенных в мягкой мозговой оболочке. Из дальнейшего изложения литературных данных, освещающих интересующий нас, вопрос, можно будеть видеть, насколько мало прибавили все последующие исследования к тому, что установил Иванов.
В самом деле, в целом ряде появившихся работ различные исследователи описывали отдельные нервы, проходившие вдоль сосудистой стенки, или нежные сплетения, располагавшиеся в адвентиции или на границе последней и мышечной оболочки, но в большинстве случаев не могли указать ни на природу описываемых волокон, ни на способ взаимодействия их со стенкой сосуда [Колликер (Kolliker), 1896; Оберштей-нер (Obersteiner), 1897; Губер (Huber), 1899; и др.].
Так, например, Лапинский (1913) не мог отнести наблюдавшиеся им нервные волокна на сосудах мягкой мозговой оболочки ни к разряду миэлиновых, ни к числу безмиэлиновых и считал их последними ветвями миэлиновых нервов неизвестной природы.
Разработка нового метода серебряной импрегнации нервных волокон расширила круг и предел исследований, связанных с выяснением иннервации сосудов головного мозга. Использование этого усовершенствованного метода гистологической обработки дало возможность более детального изучения периваскулярных нервов не только на сосудах мягкой мозговой оболочки, но и на сосудах самого мозгового вещества.
Как срезы, так и особенно тотальные препараты импрегнированной мягкой мозговой оболочки позволяют убедиться в большом количестве находящихся в ней аргирофильных элементов. Легкость импрегнации отдельных составных частей соединительной ткани, наряду с нервными волокнами, создает особые трудности диференцировки одних от других и может безусловно служить источником ошибок и неправильных заключений.
Нервные волокна мягкой мозговой оболочки могут быть прослежены не только на сосудах различного калибра, расположенных в ней, но и в соединительной ткани самой мягкой мозговой оболочки. Как будет видно из дальнейшего изложения, деление нервных волокон на нервы сосудов и нервы соединительной ткани является грубо схематическим, так как во многих случаях нервы сосудов участвуют в образовании сплетений в соединительной ткани и наоборот.
Согласно современным данным, на артериях и венах мягкой мозговой оболочки всех без исключения калибров вплоть до самых малых обнаруживается большое число нервов, проходящих вдоль оси сосуда или окружающих его в виде спирали. Эти нервы могут быть представлены одиночными мякотными волокнами, проходящими совершенно изолированно.
Обычно же нервы идут в виде пучков, то более, то менее толстых исостоящих частью из мякотных, частью из безмякотных волокон.
Однако, помимо такого рода смешанных нервных пучков, довольно значительное количество последних включает в свой состав только безмя-котные волокна.
При прохождении вдоль сосуда нервные пучки дают многочисленные ветви, вступающие в анастомотическую связь с такими же ветвями соседних нервных пучков.
Благодаря описанным связям, в адвентиции сосудов образуется широкопетлистое сплетение нервных волокон, тем более выраженное, чем больше диаметр сосуда, на котором оно располагается (рис. 124).
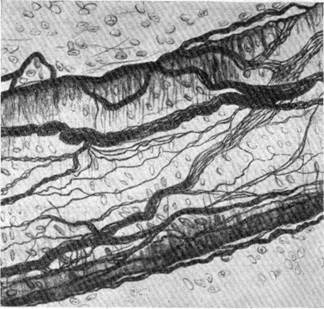
Рис. 124. Пучки нервных волокон в адвентиции кровеносного сосуда среднего калибра (по Ляховецкому).
Помимо указанного нервного сплетения, на границе адвентиции и мышечной оболочки импрегнируется второе сплетение, в состав которого входят чрезвычайно тонкие и нежные безмякотные нервы.
Согласно наблюдениям ряда авторов (Н. Д. Довгялло, 1932), это глубокое промежуточное сплетение никогда не вступает в тесные взаимоотношения с подлежащим слоем гладких мышечных волокон, так как отходящие от сплетения нервные волокна встречаются чрезвычайно редко, идут в оболочке без особого порядка и никогда не имеют концевых аппаратов.
Вместе с тем имеются и другие наблюдения, из которых следует, что от описываемого сплетения отходят волоконца, не только направляющиеся к мышечной оболочке сосуда, но и разветвляющиеся там среди гладких мышечных волокон (А. М. Ляховецкий, 1948).
Последнее обстоятельство, по мнению отдельных исследователей (А. М. Ляховецкий), указывает на моторную природу сплетения, расположенного между адвентицией и средней оболочкой сосуда. Косвенные доказательства в пользу подобного заключения основываются также на том, что промежуточное сплетение, хорошо выраженное у со-
судов с развитой мышечной оболочкой, убывает по мере уменьшения калибра сосуда, а также весьма плохо выражено у вен.
Тем не менее предположение о моторной функции нервных волокон, образующих описываемое сплетение, станет фактом лишь после того, когда будут получены убедительные данные о повсеместности отхожде-ния нервов от промежуточного сплетения в среднюю оболочку сосуда и о формировании этими нервами окончаний определенного типа среди гладких мышечных волокон.
Отсутствие до наших дней сведений относительно способа окончания нервных волокон в мышечной оболочке сосудов мягкой мозговой оболочки и неясность даже в вопросе о возможности снабжения средней оболочки нервами позволяют предполагать, что концевые образования
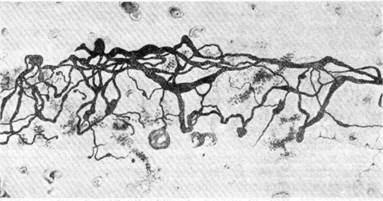
Рис. 125. Сложное нервное окончание чувствительного характера на артериоле (по Ляховецкому).
моторной природы имеют особый химический состав и не выявляются при обычных способах гистологической обработки, принятых в настоящее время.
В противоположность еще не описанным нервным окончаниям моторной природы в мышечной оболочке, концевые аппараты чувствительного характера с легкостью обнаруживаются импрегнационными методами. Чувствительные окончания самого разнообразного строения располагаются в адвентиции сосудов мягкой мозговой оболочки и представляют собой концевые аппараты нервных волокон двоякого рода.
Они, во-первых, являются окончаниями нервных волокон, отходящих от поверхностного сплетения, расположенного на адвентиции сосуда. Во-вторых, они могут быть концевыми аппаратами нервных волокон, проходящих изолированно вне связи с нервными пучками в соединительной ткани мягкой мозговой оболочки. В этих случаях волокно, обычно среднего или крупного калибра, имеющее чрезвычайно разнообразную форму хода, при следовании по соединительной ткани дает коллатераль к встретившемуся на его пути сосуду. Окончание этого волокна всегда имеет вид чувствительного аппарата.
В ряде работ различных исследователей отмечено, что одно и то же волокно может давать чувствительные окончания в адвентиции целого ряда артерий и вен. Как можно видеть из рис. 125, наиболее выраженные концевые образования имеют весьма сложное строение и величину, достигающую 100—120 м. Такого рода чувствительные аппараты наибо-
лее часто встречаются на сосудах незначительного калибра типа арте-риол. Менее оформленные окончания чувствительной природы представляют собой кустик ветвей, на которые распадается осевой цилиндр, причем каждая из веточек оканчивается пуговками, дисками или петельками разнообразного размера.
Описанные выше одиночные нервные волокна, или стволики, включающие в свой состав незначительное количество аксонов, отдающие чувствительные ветви к сосудам мягкой мозговой оболочки, представляют собой в то же время основную систему нервных волокон самой оболочки. При своем ходе по оболочке нервные волокна идут вплотную к соединительнотканным волокнам, иногда обвивая пучки их в виде спирали. Одно и то же волокно или один и тот же стволик подобного рода может принимать участие в образовании сплетений и окончаний чувствительного характера как на сосудах, так и в ткани оболочки (П. Е. Сне-сарев, 1929; А. М. Ляховецкий, 1948, и др.).
Часто отдельное нервное волокно или пучок их при прохождении по соединительной ткани покидает ее, подходит к сосуду и дальнейший свой путь совершает вдоль стенки последнего, отдавая к адвентиции его чувствительные волокна.
Во время хода такого волокна, или стволика, вдоль сосуда оно может принимать участие в образовании поверхностного сплетения. В дальнейшем волокно покидает сосуд, продолжает свой путь в соединительной ткани, подходит к другому встретившемуся сосуду, идет вдоль последнего, вновь покидает его, проходит по соединительной ткани, где и принимает участие в образовании сплетения и дает окончания чувствительного типа на клеточных элементах соединительной ткани оболочки или среди аргирофильных и клейдающих волокон ее.
Таким образом, можно видеть, насколько приблизительным является деление нервов мягкой мозговой оболочки на нервы сосудов и нервы ткани самой оболочки.
Помимо описанных выше нервных волокон, в равной мере принадлежащих сосудам и соединительной ткани мягкой мозговой оболочки, последняя имеет собственные нервы, сплетения и большое количество чувствительных концевых аппаратов.
Сплетение собственно мягкой мозговой оболочки формируется многочисленными безмякотными нервами, природа которых еще неясна. Чувствительные окончания в большинстве своем представляют собой концевые аппараты изолированно проходящих по соединительной ткани крупных нервных волокон, часть которых, по мнению отдельных исследователей, является независимой от сосудов.
Ознакомление с литературными данными, освещающими вопросы иннервации сосудов мягкой мозговой оболочки, приводит к выводу, что гистология ее до настоящего времени не вышла еще за пределы простого описания и далека от объяснения описываемых картин.
В самом деле, во всех работах данной области, начиная с самых ранних по времени их опубликования, приводятся описания большого количества нервных волокон и пучков их, располагающихся на сосудах и в ткани мягкой мозговой оболочки.
Повсюду можно найти указание на формирование этими волокнами двоякого рода сплетений: поверхностного и более глубокого промежуточного, а также описания различных по размеру и форме многочисленных концевых чувствительных аппаратов.
Большие или меньшие подробности в описании, зависящие от техники гистологического исследования, к сожалению, не всегда или совсем не
дают морфологического объяснения фактам, полученным в физиологическом эксперименте.
Физиологическими опытами с достаточной очевидностью установлена зависимость просвета артерий мягкой мозговой оболочки и мозга от импульсов, поступающих по симпатическим и парасимпатическим волокнам. Раздражение симпатического нерва на шее сопровождается некоторым сужением артерий, раздражение VII, VIII черепномозговых нервов влечет за собой увеличение просвета артерий.
Гистологическое подтверждение фактов такого рода должно было бы заключаться в обнаружении тесной связи нервных волокон одного из описанных сплетений с мышечной оболочкой сосуда мягкой мозговой оболочки.
Однако именно по вопросу о взаимоотношениях нервных сплетений и средней оболочки существует большая неясность.
На основе одного и того же импрегнационного метода, но в разных модификациях его одни авторы не могли установить моторных окончаний в мышечной оболочке и даже наличие в ней нервных волокон отмечали лишь в редких случаях, другие же отмечали ветвление нервных волокон среди гладких мышечных волокон оболочки. Но даже исследователи, наблюдавшие проникновение тонких нервных веточек в мышечный слой сосудов мягкой мозговой оболочки, не могли отметить постоянства этого явления и заключения свои строили лишь на отдельных случаях.
Понятны стремления рассматривать сплетение, расположенное между адвентицией и мышечной оболочкой сосуда, как двигательное, что находит объяснение в самом положении нервных волокон в непосредственной близости от гладких мышц оболочки. Но непонятными кажутся утверждения некоторых гистологов (А. М. Ляховецкий, 1948), приписывающих промежуточному сплетению лишь вазоконстрикторную функцию, тогда как данные физиологического опыта указывают также и на наличие сосудорасширяющих влияний со стороны парасимпатической нервной системы. Следовательно, если признавать за промежуточным сплетением двигательную функцию, то в состав его должны быть отнесены как сосудосуживатели, как и сосудорасширители.
Опыты с раздражением симпатических и парасимпатических нервов показывают, что вазомоторный эффект с полной отчетливостью обнаруживается на артериях мягкой мозговой оболочки, диаметр которых превышает 100 м.
Гистологические данные в этом отношении разнообразны.
Одни из исследователей наблюдали нервы лишь на артериях крупнее указанного размера (например, Хоробский и Пенфильд, 1932); другие же отмечают наличие нервных сплетений на артериях всех диаметров, вплоть до мельчайших их разветвлений в мягкой мозговой оболочке, описывая их даже на венах (А. М. Ляховецкий, 1948, и др.). Более того, отдельными авторами описано присутствие промежуточного сплетения не только на мелких артериальных сосудах, но в мало развитом виде даже и на венах.
Полная симпатэктомия, а также перерезка верхнего каменистого нерва с обеих сторон, по которым, как будет видно из дальнейшего изложения, к сосудам мягкой мозговой оболочки и мозга поступают сосудорасширяющие импульсы, не ведет к полной дегенерации нервов на сосудах мягкой оболочки (Хоробский и Пенфильд, 1932). Для объяснения этого факта возможны два предположения. Первое заключается в том, что существуют и другие источники, снабжающие сосуды мягкой мозговой оболочки нервами. Можно предположить также, что, невидимому,
могут существовать перерывы по ходу нервного волокна на ганглиозных клетках и сохранившиеся волокна тогда должны быть постганглионар-ными.
В литературе неоднократно описывалось отхождение ветвей от III, VI, VIII, IX, X, XI и XII черепномозговых нервов к мягкой мозговой оболочке (Белоусов, 1899; Н. Д. Довгялло, 1932, и т. д.). Существовал взгляд, согласно которому ветви указанных выше нервов принимают участие в формировании нервных сплетений соединительной ткани мягкой оболочки, т. е. являются собственными нервами этой оболочки. Последующие исследования показали, что методами гистологического анализа чрезвычайно трудно отличать нервы собственной оболочки от нервов сосудов, поскольку отдельные нервные волокна или пучки, принадлежащие, возможно, оболочке, находятся в механической связи с нервами периваскулярных сплетений.
Очень часто, как уже указывалось, отдельные нервы или пучки их могут использовать сосуд лишь как проводник, не вступая с ним ни в какую связь (П. Е. Снесарев, 1929). Кроме того, более тщательные исследования показали, что большое количество волокон, проходящих в соединительной ткани оболочки, дают коллатерали к встречающимся на их пути сосудам или принимают участие в образовании периваскуляр-ных сплетений. При этом они отдают тонкие веточки в адвентицию сосуда, где и формируется нервное окончание чувствительного характера (Н. Д. Довгялло, 1932; Бакай (Bakay), 1941; А. М. Ляховецкий, 1948, и др.).
Таким образом, часть волокон, остающаяся на сосудах мягкой мозговой оболочки интактной после полной симпатэктомии и частичного перерыва парасимпатических путей, может быть отнесена на счет описанных выше волокон центрального происхождения.
Остается рассмотреть литературные данные о наличии в сплетениях ганглиозных клеток, на которых был бы возможен перерыв преганглио-нарного волокна.
Имеется ряд указаний, согласно которым по ходу нервных сплетений, располагающихся на сосудах мягкой мозговой оболочки, находятся униполярные нервные клетки различной формы (Н. Д. Довгялло, 1932; Хоробский, 1932).
Однако в работе, опубликованной в 1948 г. и вышедшей из такой авторитетной гистологической лаборатории, какой является лаборатория Б. И. Лаврентьева, отрицается наличие нервных клеток в сплетении. Формы, напоминающие ганглиозные клетки, рассматриваются как патологическое изменение осевых цилиндров (А. М. Ляховецкий, 1948).
Отсутствие единогласия в данной области оставляет открытым вопрос о способе прохождения волокон для сосудов мягкой мозговой оболочки через симпатические и парасимпатические ганглии. В самом деле, остается невыясненным, все ли волокна симпатической природы прерываются, проходя через верхний шейный или звездчатые узлы и идут к мозгу в виде безмиэлиновых постганглионарных волокон, или же часть их идет через узлы не прерываясь, проходит известный путь по сосуду как миэлиновое преганглионарное волокно и становится безмиэлиновым только после перерыва на нервной клетке, расположенной на самом сосуде.
То же может быть сказано и в отношении хода парасимпатических волокон.
Несмотря на целый ряд попыток установить ход чувствительных нервных волокон от афферентных концевых аппаратов в адвентиции
сосудов мягкой мозговой оболочки, проблема чувствительных путей от мозговых сосудов не решена до настоящего времени и требует дальнейших исследований.
Сделанные в этом отношении заявления не выходят за пределы простых предположений и не подкреплены пока фактическими данными. Так, неоднократно поднимался вопрос о прохождении чувствительных путей через ветви блуждающего нерва, через верхний каменистый нерв, по ветвям от тройничного нерва. Однако представленные описания вызывают возражения и не могут считаться прочно установленными.
Как следует из краткого обзора состояния вопроса об иннервации сосудов мягкой мозговой оболочки, в настоящее время многое остается еще очень мало разработанным. Мы считаем, что только сочетание физиологического эксперимента с перерезкой нервных путей к сосудам мозга с тщательным гистологическим анализом результатов этих опытов может пролить свет на не решенные до сего дня вопросы о нервной ре-гуляции тока крови в мозгу.
Несмотря на то, что учение об иннервации внутримозговых сосудов находится еще на сравнительно ранней стадии своего развития, в боль-шинстве вопросов наблюдается однозначность результатов, полученных различными исследователями.
Лишь немногие, в том числе Бергер (Berger, 1924), и Хэссин (Has-sin, 1929), отрицают существование нервных волокон на сосудах мозгового вещества, у большинства же авторов наличие их не вызывает никакого сомнения.
Современные методы серебряной импрегнации позволяют убедиться в том, что внутримозговые сосуды человека, обезьяны, кошки, собаки, морской свинки, белой мыши, летучей мыши и т. д. сопровождаются нервными волокнами.
Положение о наличии периваскулярных нервов на внутримозговых сосудах в равной мере относится ко всем отделам головного мозга: различным долям полушарий, подкорковым ядрам, мозжечку, продолговатому мозгу, спинному мозгу, а также к сосудам как коры, так и белого вещества (Т. А. Григорьева, 1932; Пенфильд, 1932; Хоробский, 1932; Гумфрей (Hymphreus), 1939, и т. д.).
Нервные волокна сосудов мозга представляют собой продолжение нервных сплетений мягкой мозговой оболочки и вступают в мозг вместе с сосудами. В зависимости от диаметра сосуда, количество обнаруживаемых волокон на нем будет различным. Так, по данным Гумфрей (1939), сосуды диаметром в 250—225 мимеют приблизительно 86 нервных волокон, при диаметре сосуда в 150—125 мнасчитывается уже только 34 волокна, на сосудах диаметром 75—50 м—5 волокон, а на сосудах диаметром 25 и — всего одно волокно. Диаметр интрацеребральных сосудистых нервов очень невелик. Большинство из них имеет в поперечнике ют 0,5 до 1 м. диаметр наиболее толстых волокон не превышает 2,5 м (Пенфильд, 1932).
Периваскулярные нервы, вступив в мозг вместе с сосудом, идут или вдоль его длинной оси, или же окружают сосуд неправильными спиралями. Иногда по ходу их отходит волокно или группа их, прежде чем сам ствол разделится. Между соседними стволами могут наблюдаться довольно сложные взаимоотношения. Часть волокон одного из стволов иногда переходит в состав другого и обратно, благодаря чему стволы обмениваются волокнами. Ствол при прохождении вдоль сосуда дает две ветви, которые делятся снова и таким образом ветвление продолжается вплоть до того, пока ствол не уменьшится настолько, что дальнейшее
ветвление его невозможно. Описано также прохождение нервных волоков» по сосуду без ветвления в виде тонкой нити (Пенфильд, 1932).
Боковые ветви основных стволов с их многочисленными разветвлениями формируют тонкое сплетение нервных волокон, расположенное в адвентиции. От этого сплетения, по сообщению Гумфрей, отходят веточки, служащие для образования внутреннего сплетения. Внутреннее сплетение, расположенное непосредственно над мышечной оболочкой сосуда, состоит из тончайших нервных волокон и их разветвлений. От этого сплетения в среднюю оболочку сосуда проникают тонкие волокна, по одним-данным, имеющие оформленные нервные окончания в виде кисточек винограда диаметром, не превышающим 5 м, по другим—в виде свободных окончаний (Пенфильд, 1932; Гумфрей, 1939). Иногда такого рода-окончания обнаруживаются непосредственно на мышечной оболочке (Хоробский, 1932).
Что касается природы описываемых нервов, то большинство исследователей (Т. А. Григорьева, Пенфильд, Хоробский, Гумфрей) склонны рассматривать их как вазомоторные, частью принадлежащие симпатическому, частью парасимпатическому отделу вегетативной нервной системы.
В пользу такой точки зрения говорит структура нервных волокон, представляющих собой безмиэлиновые нервы, покрытые ядерной шван-новской оболочкой. Характерно, что интрацеребральные нервы могут являться продолжением не только безмякотных волокон сплетений мягкой мозговой оболочки, но продолжением толстых мякотных волокон, теряющих свой миэлин при входе в вещество мозга и распадающихся на тонкие нежные волокна, одетые шванновской оболочкой с ядрами (Пенфильд) .
В литературе имеются указания и на наличие на артериях внутри мозга другой группы волокон, которые, по всей вероятности, должны быть отнесены к категории чувствительных [Курузу и Хамада (Kurusu-а. Hamada), 1929; Бакай, 1941]. Однако большинством исследователей эта группа волокон обнаружена не была.
Наименее разработанной стороной учения об иннервации сосудов. мозга является вопрос о нервном снабжении капилляров.
Специальные исследования, предпринятые с целью изучения стенок мозговых сосудов, показали, что последние обладают определенной мышечной оболочкой. Значительно более тонкая, чем в сосудах других участков тела, эта оболочка тем не менее формируется из гладких мышечных волокон с овальными ядрами и гранулярной протоплазмой.
Артерии, диаметр которых превышает 25 ft, имеют двухслойную среднюю оболочку, состоящую из слоя циркулярных и слоя продольных волокон. У артерий меньшего диаметра наблюдаются лишь продольные мышечные волокна, которые и могут быть прослежены вплоть до арте-риол диаметром в 10 м.
В соответствии с этим некоторые исследователи смогли обнаружить, нервное сопровождение у артерий не менее 10 в диаметре (Гумфрей), но на сосудах меньшего диаметра нервы удалось наблюдать лишь Т. А. Григорьевой (1932).
На серебряных препаратах, обработанных по методу Гольджи, Т. А. Григорьева проследила отчетливо выраженные нервные волокна, не только сопровождавшие капилляр, но и вступавшие со стенкой его-в тесные взаимоотношения с помощью своеобразных окончаний, имевших-форму кнопок. Такого рода нервные волокна встречались даже на спавшихся капиллярах и часто имели окончание в виде кнопочки.
Незначительное количество наблюдений, посвященных специальному исследованию наличия нервов на капиллярах мозга, не дает возможности нидля подтверждения, ни для отрицания фактов, сообщенных Т. А. Григорьевой.
Трудность гистологического анализа среди массы аргирофильных элементов мозга заставляет отдельных авторов склоняться к тому, что их неудачи в обнаружении нервов на капиллярах должны быть отнесены за счет недостаточного совершенства употреблявшегося ими метода обработки (Пенфильд, 1932).
ГлаваIX
Дата добавления: 2015-05-16; просмотров: 1161;