ДЕЙСТВИЕ
Наиболее важным биологическим эффектом вазопрессина является сохранение воды в организме путем снижения выделения мочи. Этот антидиуретический эффект осуществляется за счет усиления реабсорбции не содержащей растворенных веществ в дистальных и/или собирательных канальцах почки [36] (рис. 9—7). В отсутствие вазопрессина мембраны, выстилающие эту часть нефрона, формируют непреодолимое препятствие для диффузии воды и растворенных веществ. Гипотонический фильтрат, образующийся в более проксимальных отделах нефрона, без изменения проходит через дистальный каналец и собирательный проток. В таких условиях, называемых водным диурезом, осмоляльность и ток мочи у здорового взрослого человека обычно составляют примерно 60—70 мосммоль/кг и 15—20 мл/мин соответственно.
В присутствии вазопрессина гидроосмотическая проницаемость дистальных и собирательных канальцев увеличивается, что создает возможность диффузии воды по осмотическому градиенту,. существующему в норме между канальцевой жидкостью и изотонической или гипертонической средой коркового и мозгового слоя почки (см. рис. 9—7). Поскольку вода реабсорбируется без растворенных веществ, осмотическое давление оставшейся мочи повышается, а ее объем и скорость тока уменьшаются. Количество воды, реабсорбируемой в дистальных отделах нефрона, а следовательно, и степень концентрированности мочи изменяются как функция концентрации вазопрессина в плазме (рис. 9—8). У здорового взрослого человека эта зависимость между стимулом и реакцией крайне чувствительна, поскольку все возможные значения концентрированности и разведения мочи определяются изменением уровня вазопрессина в плазме всего в 10 раз или менее. При максимально эффективном содержании вазопрессина осмоляльность и скорость тока мочи у человека составляют приблизительно 1200—1400 мосммоль/кг и 0,3—0,6 мл/мин соответственно. У некоторых животных, особенно у грызунов, моча может быть намного более концентрированной, по-видимому, за счет более длинных почечных сосочков и соответственно более высокой гипертоничности в мозговом слое.
Влияние вазопрессина на концентрированность и/или скорость тока мочи в значительной мере зависит от изменения объема фильтрата, находящегося в дистальных канальцах. У здорового взрослого человека 85—90% примерно из 200 л плазмы, фильтрующейся за сутки в клубочках, реабсорбируется изоосмотически с натрием и глюкозой в проксимальной части нефрона. Остающиеся 20 л становятся гипотоничными вследствие избирательной реабсорбции натрия в восходящем колене петли Генле и достигают дистальной части нефрона, где в зависимости от активности вазопрессина может избирательно реабсорбироваться еще до 19 л/сут. При высоком потреблении соли и повышенном количестве плохо реабсорбирующихся растворенных веществ, подобных
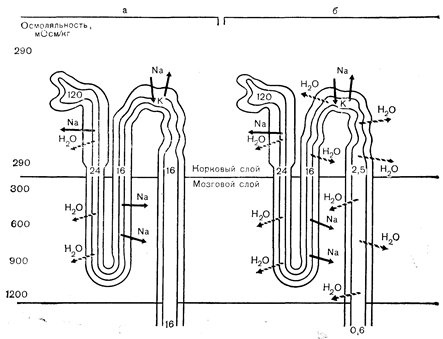
Рис. 9—7. Схематическое изображение влияния вазопрессина (ВП) на образование мочи в нефроне. Осмотическое давление ткани и канальцевой жидкости показано плотностью затемнения фона. Цифры в просвете нефрона обозначают типичные скорости тока мочи (мл/мин). Стрелки указывают на реабсорбцию натрия (Na) или воды (Н2О) с помощью активного (сплошные) или пассивного (пунктирные) механизма. Можно видеть, что ВП действует только на дистальную часть нефрона, где он увеличивает гидроосмотическую проницаемость канальцевых мембран. Жидкость, достигающая этой части нефрона, в норме составляет 10—15% от общего фильтрата и гипоосмотична благодаря избирательной реабсорбции натрия в восходящем колене петли Генле. В отсутствии ВП (а) мембраны дистальной части нефрона остаются сравнительно непроницаемыми для воды и растворенных веществ, и жидкость, проходящая через петлю Генле, выделяется в виде мочи в основном без изменений. В условиях максимального эффекта ВП (б) большая часть воды (за исключением 5—10%) из этой жидкости пассивно реабсорбируется в норме по осмотическому градиенту, существующему между ней и окружающей тканью. маннитолу, мочевине или глюкозе, проксимальный каналец может пропускать больше 10—15% фильтрата. В результате объем жидкости, достигающей дистальной части нефрона, может превысить ограниченную способность последнего к реабсорбции воды. Вследствие этого осмоляльность мочи снижается, а скорость ее тока увеличивается даже в присутствии повышенного уровня вазопрессина. Этот тип полиурии называют диурезом растворенных веществ, чтобы отличить его от диуреза, обусловленного недостаточностью эффекта вазопрессина. Наоборот, при некоторых клинических состояниях, например, застойной сердечной недоста-
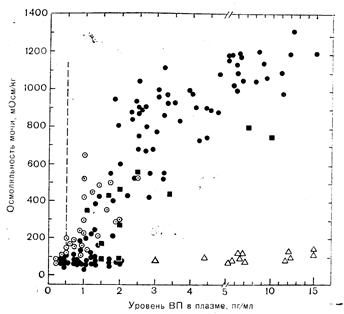
Рис. 9—8. Зависимость осмоляльности мочи от концентрации вазопрессина (ВП) в плазме у здоровых взрослых лиц, находящихся в состоянии гидратации, и у больных с полиурией различной этиологии.
Черные кружки — здоровые лица (23 человека); черные квадраты — 2 больных с первичной полидипсией; светлые треугольники — 2 больных нефрогенным несахарным диабетом: кружки с точкой — 8 больных гипофизарным несаха,рным диабетом. точности, при которых в проксимальной части нефрона реабсорбируется повышенное количество фильтрата, способность экскретировать не содержащую растворенных веществ воду резко снижается даже в отсутствие эффекта вазопрессина.
Антидиуретический эффект вазопрессина может ингибироваться и в условиях уменьшения концентрационного градиента в мозговом слое почек. Это может иметь столь разные причины, как хронический водный диурез, снижение кровотока в мозговом слое почек или белковая недостаточность. Однако в силу того, вероятно, что в дистальных отделах извитого канальца и/или проксимальной части собирательного протока может изотонически реабсорбироваться большой объем жидкости, вытекающей из петли Генле, уменьшение лишь концентрационного градиента в мозговом слое почек редко приводит к значительной полиурии.
Клеточный механизм, с помощью которого вазопрессин изменяет гидроосмотическую проницаемость эпителия дистальных канальцев, выяснен не полностью. Однако имеются многочисленные данные, свидетельствующие о том, что гормон связывается рецепторами серозной поверхности клетки, что в свою очередь активирует аденилатциклазу и тем самым увеличивает гидроосмотическую проницаемость противоположной, или слизистой, поверхности клетки [37]. Характер и последовательность биофизических процессов, «запускаемых» цАМФ, во многом остаются областью предположений, но они должны были бы включать открытие высокоспециализированных каналов или пор при участии клеточных микротрубочек [37, 38]. Имеются также некоторые данные, указывающие на роль простагландинов в качестве своего рода местной или «короткопетельной» системы обратной связи, модулирующей влияния вазопрессина на почки [39]. Так, вазопрессин стимулирует синтез простагландина Е, который в свою очередь ингибирует влияние гормона на аденилатциклазу и гидроосмотический ток. Напротив, ингибиторы синтеза простагландинов, такие, как индометацин, потенцируют антидиуретический эффект гормона. Синтез простагландинов в почках стимулируется также ангиотензином, брадикинином и гипертонической средой. Это позволяет думать о том, что влияние различных факторов на экскрецию мочи может опосредоваться изменением почечного эффекта вазопрессина.
Вазопрессин действует также на различные внепочечные системы, такие, как кровеносные сосуды [40], желудочно-кишечный тракт [41] и ЦНС [42]. Концентрация вазопрессина, необходимая для проявления его внепочечных эффектов, точно не известна, но должна была бы, по-видимому, быть во много раз большей, чем та, в которой он содержится в плазме периферической крови в физиологических условиях. Тем не менее не исключено, что при резком повышении секреции вазопрессина, вызываемом такими стимулами, как гипотензия или тошнота, его уровень повышается в достаточной степени, чтобы повлиять на сосудистый тонус, сокращение желудочно-кишечного тракта или воротный кровоток. Клеточные механизмы внепочечных эффектов вазопрессина совершенно не известны. Однако на сосуды и почечные канальцы действуют разные части молекулы гормона, поскольку структурные модификации, которые приводят к потере прессорной активности» не только не уменьшают, но даже увеличивают антидиуретическую активность его [6, 7].
В отношении влияния вазопрессина на переднюю долю гипофиза и/или гипоталамуса все еще существуют разногласия [43]. Хотя введение относительно больших доз вазопрессина в общий кровоток стимулирует секрецию АКТГ и гормона роста, большинство данных свидетельствует, по-видимому, о том, что эти эффекты не являются прямыми. Эндогенная секреция вазопрессина наверняка не обязательна для активации гипофизарно-адреналовой системы, поскольку у крыс линии Brattleboro, лишенных способности синтезировать этот гормон, практически полностью сохраняется реакция на различные формы стресса.
Основной биологический эффект окситоцина заключается в облегчении грудного кормления, так как он вызывает выброс молока. Это происходит путем сокращения миоэпителиальных клеток лактирующей молочной железы. Окситоцин может также способствовать родам, стимулируя сокращение матки. Оказывает ли гормон какое-либо существенное влияние у мужчин — неиз- вестно. При экзогенном введении он может вызывать легкий диурез и/или натрийурез, но не известно, достигает ли когда-либо содержание эндогенного гормона в крови того уровня, при котором проявляется этот эффект.
Биологические эффекты нейрофизинов не установлены за тем исключением, что они образуют комплексы с окситоцином и вазопрессином в нейросекреторных гранулах нейрогипофиза. Хотя они и присутствуют в плазме, но не играют роли связывающих или транспортных белков, так как более высокое значение рН и низкая концентрация лигандов обусловливают полную диссоциацию комплексов.
ЖАЖДА
У наземных животных механизм жажды служит незаменимым дополнением антидиуретической регуляции водного баланса. В физиологическом смысле термин «жажда» используют для обозначения осознанного ощущения потребности в воде. Жажду следует отличать от других детерминант потребления воды, таких, как социальные привычки или обычная диета. Как и следовало ожидать, жажда стимулируется многими факторами, вызывающими секрецию вазопрессина [44]. Наиболее эффективным из них является, по-видимому, гипертоническая среда. У здоровых взрослых лиц повышение осмоляльности плазмы всего на 2—3% по сравнению с исходным уровнем приводит к сильному чувству жажды. Эта реакция не зависит от изменений внеклеточного объема жидкости, поскольку она сохраняется при повышении осмоляльности плазмы как путем ограничения приема жидкости, так и путем инфузии гипертонического солевого раствора. Абсолютный уровень осмоляльности плазмы, при котором появляется жажда, можно назвать осмотическим порогом жажды. Он несколько колеблется у разных индивидуумов, но в среднем в популяции здоровых взрослых лиц составляет около 295 мосмоль/кг [16]. Этот уровень существенно выше осмотического порога секреции вазопрессина и близко совпадает с тем, при котором в норме достигается максимальная концентрированность мочи (см. рис. 9—3 и 9—8). Таким образом, механизм появления жажды можно считать своего рода тормозом, главная функция которого заключается в том, чтобы не допустить развития или корригировать ту степень дегидратации, которая превышает компенсаторные возможности антидиуретической системы.
Нервные пути, опосредующие появление жажды, выяснены недостаточно. Однако они должны были бы, по-видимому, включать осморецептор, расположенный в переднебоковом гипоталамусе вблизи к осморецептору, регулирующему секрецию вазопрессина, но не целиком совпадающему с ним. Оба рецептора обладают, по-видимому, сходной специфичностью, так как те растворенные в плазме вещества (например, мочевина или глюкоза), которые практически не влияют на секрецию вазопрессина, в равной мере неэффективны и в отношении стимуляции жажды. Чувствительность обоих осморецепторов к более эффективным растворенным веществам, таким, как натрий, не поддается строгому сравнению, но, вероятно, тоже близка. Чувство жажды быстро усиливается в прямой зависимости от осмоляльности плазмы и обычно становится почти невыносимым, когда осмоляльность всего на 10—15 мосмоль/кг превышает пороговый уровень. Потребление воды, по-видимому, пропорционально интенсивности чувства жажды и в условиях длительной или максимальной стимуляции может достигать примерно 20—25 л/сут.
Уменьшение объема или давления крови также вызывает жажду. У человека степень гиповолемии и/или гипотензии, необходимая для того, чтобы вызвать чувство жажды, не определена, но должна превышать ту, которая влияет на секрецию вазопрессина. Таким образом, маловероятно, чтобы гемодинамические параметры оказывали существенное влияние на потребление воды, кроме как в патологических условиях. Механизм, с помощью которого гиповолемия и/или гипотензия вызывают жажду, неясен,. но может заключаться в «перенастройке» осморегуляторной системы подобно тому, что показано в отношении вазопрессина. Эти эффекты опосредуются, вероятно, теми же волюм- и барорецепторными механизмами, но могут зависеть (по крайней мере отчасти) и от системы ренин — ангиотензин, которая у животных является мощным дипсогенным фактором, хотя у человека аналогичные эффекты пока не показаны.
Жажду у человека вызывает также введение 2-дезоксиглюкозы. Однако инсулиновая гипогликемия является намного менее дипсогенным фактором, хотя секрецию вазопрессина она стимулирует в той же степени, что и 2-дезоксиглюкоза. Такая диссоциация указывает на то, что эти два фактора (по крайней мере частично) действуют различными механизмами.
Дата добавления: 2015-01-19; просмотров: 1473;