Исследования нервной проводимости
Стимулирование достаточно больших моторных и чувствительных нервов позволяет регистрировать их потенциалы действия и получать объективные количественные данные относительно латентного периода и скорости проведения импульса по нерву. Методика основана на стимулировании нерва поверхностными электродами, помещаемыми на кожу над исследуемым нервом. В результате электроды регистрируют составной потенциал действия в нерве, расположенном проксимально (при исследовании больших сенсорных волокон) или над мышцей, расположенной дистально (в случае исследования моторных волокон в смешанном двигательно-чувствительном нерве) (рис. 354-3). Время проведения импульса от наиболее дистально расположенного стимулируемого электрода, измеренное в миллисекундах, с момента стимуляционного воздействия до начала ответной реакции, получило название дистального, или периферического латентного, времени. Если второй стимул наносится на смешанный нерв более проксимально (или если регистрирующие электроды расположены более проксимально в случае исследования чувствительных волокон), то можно измерить новое и более продолжительное время проведения. Если расстояние (в миллиметрах) между двумя участками стимуляции двигательных волокон или регистрации с чувствительных волокон разделить на разницу показателей времени проведения (в миллисекундах), можно получить максимальную скорость проведения (в метрах в секунду). Она показывает скорость распространения потенциалов действия в наиболее крупных и наиболее быстро проводящих импульс нервных волокнах. Показатели этих скоростей у здоровых лиц колеблются довольно значительно — от 40—45 м/с (в зависимости от исследуемого нерва) до 75—80 м/с. У новорожденный эти значения несколько ниже (они составляют примерно 1/2 от показателей взрослых), но достигают данного уровня к 3—4 годам жизни. Получены показатели нормы периферических латентных периодов для наиболее дистальных участков различных смешанных нервов, иннервирующих соответствующие мышцы. Например, когда стимулируют срединный нерв у запястья, латентный период для проведения импульса через канал запястья к короткой отводящей мышце большого пальца у здоровых лиц составляет менее 4,5 м/с. Составлены специальные таблицы с подобными нормативами для скорости проведения и дистальных латентных периодов, показатели которых колеблются в зависимости от расстояния. Во время исследования нервной проводимости очень важно поддерживать нормальную температуру тела человека, так как при субнормальной температуре скорость проведения по нерву замедляется. Скорость проведения по нерву зависит от диаметра нервного волокна и степени его демиелинизации. Немиелинизированные нервные волокна малого диаметра характеризуются меньшей скоростью проведения, чем миелинизированные волокна большого диаметра. У волокон с сегментарной демиелинизацией скорость проведения, как правило, уменьшена. При стимулировании моторных волокон периферического нерва при условии, что каждое нервное волокно находится в функциональном единении с иннервируемыми им многочисленными мышечными волокнами, с кожного электрода над исследуемой мышцей можно зарегистрировать составной мышечный потенциал действия, являющийся результатом импульсных разрядов многих мышечных волокон. Сенсорные потенциалы действия, регистрируемые непосредственно с самих нервных волокон, не обладают качеством «амплификации», создаваемым мышечными волокнами; дело в том, что для этого требуется большая электронная амплификация. В случае патологии нервов сенсорные потенциалы могут быть небольшими или их может не быть вообще, и, таким образом, зарегистрировать сенсорную проводимость становится невозможно. Напротив, достаточно надежно измерить скорость моторной проводимости возможно, даже если сохранным осталось лишь одно мышечное волокно. Измерения скорости нервной проводимости отражают
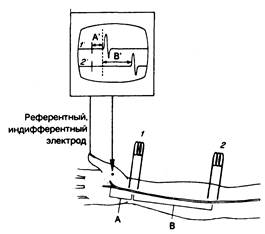
Рис. 354-3. Измерение скорости проводимости по нерву. Срединный нерв раздражают через кожу на уровне запястья (1) или в передней локтевой ямке (2), в результате чего возникает составной мышечный потенциал действия, регистрируемый как разница потенциалов между поверхностным электродом над выпуклостью тенара (вертикальная стрела) и индифферентным (референтным) электродом (РЕФ), расположенным более дистально. Изгиб кривой 1' на катодно-лучевом осциллоскопе (КЛО) отражает момент воздействия раздражения в точке 1, за которым следует мышечный потенциал. Дистальная «латентность» — это время А' на кривой КЛО (например, 3 м/с), которая соответствует проведению импульса на отрезке А по кисти.
То же самое относится и к кривой 2' (стимул воздействует в точке 2, а время от начала его воздействия до ответной реакции равно А' +В'). Максимальную скорость моторного проведения от точки 2 до точки 1 можно получить, разделив расстояние В на время В'.
состояние наиболее хорошо сохранившихся нервных волокон, и, если непораженным осталось лишь небольшое число нервных волокон, показатели нервной проводимости могут быть нормальными, несмотря на довольно распространенную нервную дегенерацию. После неполного пересечения нерва острым предметом в небольшом числе оставшихся нервных волокон максимальная скорость моторного проведения может сохраниться, хотя мышца, вовлеченная в патологический процесс, почти полностью парализована. Аксон является первичным очагом поражения при алкогольной, алиментарной, уремической и диабетической невропатиях. По оставшимся неповрежденными аксонам проведение импульсов сохранено, так что, когда поражены более массивные нервные волокна, оставшиеся интактными нервные волокна меньшего диаметра, способные нормально проводить импульс, обеспечивают слегка замедленную скорость максимального моторного проведения. При многих невропатиях скорость нервной проводимости остается нормальной или лишь незначительно снижена. Обычные исследования нервной проводимости осуществляют для того, чтобы подтвердить наличие невропатии.
При этом сравнивают результаты, полученные у испытуемых, с данными нервной проводимости в контрольной группе лиц, подобранной адекватно по возрасту и полу. Хотя многие заболевания периферических нервов не влияют на скорость нервного проведения, тем не менее такие заболевания, как острая идиопатическая полиневропатия (синдром Гийена—Барре), дифтерия, метахроматическая лейкодистрофия и гипертрофические невропатии, вызывают замедление скорости проведения, так как при этом первично поражаются шванновские клетки и наблюдается сегментарная демиелинизация. Очаговые сдавления нерва, как это бывает при синдроме ущемления нерва в костном канале, вызывают локализованное замедление проводимости вследствие сжатия аксонов и демиелинизации в участке ущемления нерва. При обнаружении такого очагового замедления нервной проводимости диагноз ущемления нерва подтверждается. Диагноз сдавления срединного нерва в канале запястья основан на сравнении периферической (терминальной) латентности (латентное время) одного срединного нерва с другим срединным нервом или с локтевым нервом. Однако даже если показатели проводимости при этом нормальны, исключить синдром ущемления нерва нельзя.
Другие методы оценки нервной проводимости. Для изучения нервной проводимости в более проксимальных сегментах осуществляют измерение латентностей для F-реакций, Н-рефлексов и мигательных рефлексов. Эти методы позволяют определить скорость проведения с периферии (конечности, лицо) к центральной нервной системе (спинной мозг или ствол мозга) и обратно. Так, F-реакция определяет время, необходимое для прохождения раздражения, нанесенного на аксон альфа-моторного нейрона, антидромно (т. е. в противоположном направлении) по направлению к переднему рогу спинного мозга и затем возвращение импульса ортодромно, обратно к тому же аксону. Н-рефлекс определяет время, необходимое для ортодромного (в прямом направлении) проведения возбуждения вверх по нерву через чувствительные волокна группы IA через спинномозговую синаптическую связь с альфа-моторным нейроном и затем ортодромно вниз к моторному аксону. Таким образом может быть измерена скорость проведения импульса по проксимальным сенсорным и моторным нервам и корешкам спинномозгового нерва. Использование указанных методик для определения скорости проведения в проксимальных нервах позволило выявить нарушение этого показателя у 80—90% больных с периферической невропатией. Мигательные рефлексы отражают скорость проведения импульсов по ветвям тройничного и лицевого нервов. Мигательный рефлекс, вызванный электрической стимуляцией супраорбитальных ветвей тройничного нерва, позволяет определить локализацию поражений в системах лицевого и тройничного нервов.
Исследования нервной проводимости, описанные выше, достаточно условны, так же как и исследования запоздалых реакций. Они информативны лишь по отношению к быстро проводящим аксонам большого диаметра, но несут мало информации о характере проведения в промежуточных нервных волокнах и волокнах небольшого диаметра. При использовании физиологических принципов сталкивания нервных импульсов, вызванных стимуляцией в двух разных областях (проксимально и дистально) одного и того же нерва, можно измерить нервную проводимость в моторных аксонах малого диаметра. Патологические скорости нервного проведения в нервных волокнах промежуточного размера наблюдали у некоторых больных с метаболическими и алиментарными невропатиями даже тогда, когда результаты обычных методов исследования нервной проводимости и F-реакции были нормальными.
Тесты с повторной стимуляцией. При патологии нервно-мышечного соединения показатели начального составного мышечного потенциала действия, вызываемого супрамаксимальным электрическим раздражением исследуемого нерва, остаются нормальными, однако после нескольких стимуляций, проводимых со скоростью 2—3 Гц, амплитуда составного мышечного потенциала действия начинает уменьшаться, но после 4—5 раздражений вновь возрастает. Такой характер снижения потенциала, достигающего максимума при 4—5-м раздражении с последующим возрастанием при продолжающихся раздражениях, характерен для миастении. Этот дефект напоминает частичную блокаду, вызываемую кураре, и отражает постсинаптическое нарушение синаптической функции. Дефект этот обратим при применении антихолинэстеразных препаратов, например при внутривенном введении эдрофониума гидрохлорида (Edrophonium hydrochloride) в дозе 5—10 мг. Прогрессирующее снижение составного мышечного потенциала действия при повторной стимуляции нерва наблюдают при полиомиелите, амиотрофическом боковом склерозе, миотонии и при другой патологии моторной единицы. Однако при этих заболеваниях не регистрируют типичную кривую уменьшения—увеличения величины потенциала, столь характерную для миастении.
При синдроме Ламберта—Итона (миастенический синдром) повторные стимуляции облегчают трансмиссию импульса. Быстрая стимуляция нерва (20—30 Гц) вызывает прогрессирующее увеличение мышечных потенциалов действия, которые очень невелики или вовсе отсутствуют поначалу, при первой стимуляции, но затем амплитуда их увеличивается до нормальных значений. Это облегчение ответной реакции не подвергается воздействию антихолинэстеразных препаратов, но может быть заторможено гуанидин-гидрохлоридом (Guanidine hydrochloride), назначаемым по 10—30 мг/кг в день дробно. Нервно-мышечный трансмиссионный дефект этого «реверсированного» миастенического синдрома является результатом патологического высвобождения ацетилхолина. Такой же дефект возникает при воздействии ботулинического токсина или при параличе, вызываемом аминогликозидными антибиотиками (см. гл. 358).
Дата добавления: 2015-03-17; просмотров: 2651;