Регуляція фотосинтезу на рівні фотосинтетичного апарату
Регуляція фотосинтезу на рівні фотосинтетичного апарату здійснюється різними механізмами. Виділяють чотири основні шляхи регуляції цього процесу.
1. Перерозподіл енергії поглинутого світла між світлозбиральними пігментними комплексами двох фотосистем.
2. Терморегуляція процесів електронного транспорту і сполучених з ним процесів переносу протонів і синтезу АТР. Процес може мати важливе значення при адаптації чутливих до холоду рослин.
3. Іонна регуляція, зумовлена перерозподілом катіонів ( у т.ч. протонів) та аніонів між стромою і внутрішньотилакоїдним простором хлоропластів.
4. Редокс-залежна регуляція (зміна окисно-відновного стану переносників ланцюга електронного транспорту або окремих метаболітів).
6.8. Фотосинтез і обмін речовин у рослинній клітині
Цикл відновлення СО2 до цукрів повністю відбувається у хлоропластах. За кількістю синтезованого продукту головною є сахароза. Складовими для її утворення є перші відновлені продукти фотосинтезу ФГА і ДОАФ, які легко транспортуються через мембрани хлоропластів. Поряд з цим частина фосфотріоз окисляється і дефосфорилюється, стаючи джерелом енергії (АТР і НАДР.Н) і субстратом для різних біосинтетичних реакцій клітини. Тому поняття “продукт фотосинтезу” поширюється не тільки на проміжні й кінцеві вуглеводи, що синтезуються у циклі Кальвіна у хлоропластах, але й на сполуки, що утворилися на основі цих проміжних сполук за участю енергії АТР і НАДР.Н, створених у світловій стадії фотосинтезу (рис. 39).
 Рис. 39. Використання проміжних продуктів циклу Кальвіна на синтез різних сполук хлоропластів і клітини: РБФ – рибулозобіфосфат; ФГК – фосфогліцеринова кислота; ФГА – фосфогліцериновий альдегід; Ф6Ф – фруктозо-6-фосфат; ФЕП – фосфоенолпіровиноградна кислота; КоА – коензим-А; ЩОК – щавлевооцтова кислота.
Рис. 39. Використання проміжних продуктів циклу Кальвіна на синтез різних сполук хлоропластів і клітини: РБФ – рибулозобіфосфат; ФГК – фосфогліцеринова кислота; ФГА – фосфогліцериновий альдегід; Ф6Ф – фруктозо-6-фосфат; ФЕП – фосфоенолпіровиноградна кислота; КоА – коензим-А; ЩОК – щавлевооцтова кислота.
Близько половини усієї кількості АТР, яка накопичується при фотосинтетичному фосфорилюванні, використовується на процес асиміляції СО2 у вуглецевих циклах і утилізується у кінцевих продуктах фотосинтезу. Решта (40–50%) витрачається на активацію метаболічних систем і конструктивних процесів у рослині, на підтримку структур фотосинтетичного апарату, відновні та інші процеси життєдіяльності клітини.
6.8.1. Біосинтез полісахаридів, амінокислот, білків,
ліпідів, фітогормонів, полісахаридів
Сахароза і крохмаль – це одні з перших нефосфорильованих продуктів фотосинтезу. Близько 50–70 % вуглецю, засвоєного при фотосинтезі, використовується на утворення сахарози. Її біосинтез збільшується при посиленні використання асимілятів. Якщо відтік з листків цієї основної транспортної форми уповільнюється, то її концентрація підвищується. Одночасно відбувається нагромадження тріозофосфатів і гексозофосфатів, які пригнічують активність ферменту сахарозосинтетази, а отже – і синтез сахарози. Пригнічення синтезу сахарози призводить до накопичення у хлоропластах асимілятів, з яких синтезується крохмаль. Асиміляційному крохмалю належить роль тимчасового запасного продукту, що використовується як джерело енергії і пластичних речовин у темний період доби. Він відіграє також буферну роль у стабілізації процесу фотосинтезу. Ще одна функція крохмалю полягає у забезпеченні й підтриманні фонду проміжних сполук фотосинтетичного циклу відновлення вуглецю, оскільки при його гідролізі утворюється глюкозо-1-фосфат, здатний перетворюватися у фруктозо-6-фосфат. Підвищення кількості крохмалю у хлоропластах найчастіше є результатом погіршення експорту продуктів фотосинтезу від хлоропластів до цитоплазми.
Його збільшення серед продуктів фотосинтезу також відбувається з поглибленням диференціації хлоропластів та їх старінням.
Значна частина продуктів фотосинтезу використовується у процесах асиміляції азоту і синтезу азотовмісних сполук. Численні дослідження свідчать про зв’язок вуглецевого й азотного обміну, і тому на сьогодні хлоропласти розглядаються як центри сполучення цих процесів, які є основою продуктивності рослини.
При посиленні азотного живлення помітно зростає споживання синтезованих асимілятів на внутрішньоклітинні синтетичні процеси.
З кінцевих азотовмісних сполук найбільше значення мають білки і фотосинтетичні пігменти. Безпосереднім попередником структурних елементів клітини (білків) є амінокислоти. Синтез і співвідношення амінокислот у листках залежать від чергування світла і темряви протягом вегетації і є генотипною ознакою.
6.9. Транспортування метаболітів
6.9.1. Транспорт асимілятів і його регуляція у листковій пластинці
Транспортну роль у листковій пластинці виконує система з чисельних жилок. Рух речовин у листку відбувається по ксилемі і флоемі. Основними провідними елементами ксилеми всіх жилок, за винятком кінцевих, є судини (трахеї). Найменші кінцеві жилки представлені трахеїдами – неперфорованими клітинами, що мають пори на загальних стінках.
Провідними елементами флоеми є ситоподібні трубки і клітини-супутниці. Особливостями будови ситоподібних трубок є відсутність ядра і тонопласту, наявність густої розгалуженої сітки фібрил і дуже обводненої протоплазми. Ці особливості зумовлюють їх спеціалізацію до активного транспорту речовин.
Флоемна частина кінцевих відгалужень жилок у листку складається з паренхімних клітин.
Основною ознакою провідної системи листка є її зв’язок з мезофілом. Між мезофілом і провідною системою знаходиться вільний простір. Очевидно, за допомогою сітки провідних пучків можлива адсорбція цукрів та інших асимілятів з вільного простору, концентрація їх у флоемі для відведення з листка. Продовженням провідної системи листкової пластинки є провідна система черешка.
У листку існують два шляхи транспорту речовин: дальній – по флоемі і ксилемі, який забезпечує координацію функцій усієї рослини, і внутрішньоклітинний (ближній), який забезпечує пересування речовин у клітині. Останній здійснюється, в основному, за допомогою цитоплазматичних мембран. Обидва шляхи транспорту функціонують одночасно і взаємнопов’язано.
У найбільш типових випадках транспорт метаболітів починається у фотосинтезуючих хлоропластах, з яких він поширюється до цитоплазми, потім проходить через низку паренхімних клітин і, подолавши концентраційний поріг, продовжується у системі клітин флоеми до споживаючих органів. Тут метаболіти знову переходять на етап паренхімного транспорту і закінчують пересування у запасаючих або споживаючих клітинах.
6.9.2. Паренхімний транспорт асимілятів
У листку відбувається насамперед ближній транспорт пластичних речовин, який проходить через систему симпластів.
Відтік асимілятів на короткі відстані (ближній транспорт) може бути зумовлений виникненням і дією осмотичних факторів, що сприяють переміщенню продуктів від клітин з високим осмотичним тиском по градієнту концентрації. Якщо оболонка фотосинтезуючої клітини тоненька, то цукри можуть переміщуватись осмотичним шляхом. При наявності у клітинних стінках поясків Каспарі транспорт продуктів фотосинтезу здійснюється симпластним шляхом, якому сприяють численні плазмодесми.
Розпочавши свій шлях у фотосинтезуючих хлоропластах, метаболіти розповсюджуються по цитоплазмі й досягають клітинної стінки. У вільному просторі може нагромаджуватися до 20% загальної кількості синтезованих листками вуглеводів. Основним продуктом фотосинтезу у клітинах губчастої паренхіми є сахароза, а у полісадній паренхімі синтезується головним чином крохмаль. Вважають, що губчаста паренхіма більшою мірою, ніж палісадна, спеціалізована до транспорту асимілятів. Саме до губчастої паренхіми повернута добре розвинена флоема. Заповнення флоеми продуктами фотосинтезу розпочинається з найменших елементів провідної системи. Вільний простір є проміжною зоною у відтоці асимілятів від фотосинтезуючих до провідних клітин листкової пластинки.
Транспорт продуктів фотосинтезу у листку залежить від інтенсивності фотосинтезу, онтогенетичного розвитку листка і рослинного організму в цілому. При мінімальній експортній функції листка більше утворюється вуглеводних продуктів, які витрачаються на побудову власних структурних елементів клітини. З ростом листка і формуванням його роль, як джерела асимілятів для тканин і інших органів, зростає. Збільшується загальна і відносна інтенсивність утворення сахарози. При досягненні остаточного розміру листка утворення сахарози стає домінуючим процесом.
Оскільки у молодого листка інтенсивність фотосинтезу значно нижча ніж у дорослого, його біосинтетичні процеси з утворення нуклеїнових кислот, білків, ліпідів та інших складних сполук не можуть бути забезпечені тільки власними асимілятами, тому відбувається приплив асимілятів з інших органів. За таких умов власні асиміляти практично не досягають транспортних систем листка, і здійснюється тільки ближній транспорт.
Іншою причиною слабкого транспорту фотосинтетичних продуктів від молодих листків може бути і відсутність потужного споживача.
6.9.3. Флоемний транспорт фотоасимілятів
Далекий транспорт пластичних речовин здійснюється по флоемі, яка складається із ситоподібних трубок і клітин-супутниць. Ситоподібні трубки є системою клітин, поперечні стінки яких пронизані розширеними плазмодесмами. Ці клітини не мають тонопласта. У них добре розвинена внутрішня поверхня, представлена тонкими нитками цитоплазми, що, очевидно, є важливим для транспортної функції. До ситоподібних трубок прилягають клітини-супутниці, сполучені з елементами ситоподібних трубок численними плазмодесмами.
Асиміляти від місця їх утворення транспортуються симпластним шляхом до клітин-супутниць і паренхімних клітин, оточуючих ситоподібні трубки. Цей транспорт потребує витрат енергії, тому процес гальмується відсутністю кисню, зниженням температури та ін.
При наявності у клітині фосфатів глюкоза перетворюється у сахарозу й активно транспортується до ситоподібних трубок. У місцях споживання відбувається вихід сахарози із ситоподібних трубок у формі глюкози і рух симпластним шляхом до окремих клітин споживаючих органів. Швидкість пересування асимілятів – 50–100 см/год.
Інтенсивність транспорту асимілятів визначається потребою у них органів і частин рослин, що формуються. Транспорт по рослині має чітку спрямованість. Нижні листки забезпечують асимілятами кореневу систему, а верхні – висхідний потік. Крім того, у рослині наявний і поперечний транспорт у тканинах без власної провідної системи. Розподіл асимілятів по рослині має низку обмежень, пов’язаних з морфологією та ендогенною регуляцією цього процесу.
Пересування органічних речовин у рослині по ситоподібних трубках флоеми проходить відносно швидко на великі відстані завдяки численним плазмодесмам, що утворюють симпластну систему. Відтік асимілятів від листків включає три взаємопов’язаних процеси: завантаження флоеми шляхом виштовхування продуктів фотосинтезу з асимілюючих клітин, транспорт по елементах флоеми і розвантаження у місцях їх споживання.
Механізм перенесення і накопичення органічних речовин залежить від активної функції ситоподібних трубок і клітин-супутниць, яка зумовлюється гідрофільністю колоїдів, диханням, обмінною адсорбцією іонів.
Завантаження флоем цукрами у провідній системі листка проходить проти градієнта концентрації і супроводжується витратою енергії, джерелом якої є процеси дихання у клітинах-супутницях. Встановлена залежність між диханням і накопиченням у клітинах розчинених речовин.
Згідно з сучасними уявленнями, у плазмалемі клітин-супутниць і паренхімних клітин флоемних закінчень тонких жилок існує водневий насос, що працює на зовнішній бік мембрани. Трансмембранне перенесення іонів водню Н+ у симпорті із сахарозою здійснюється за допомогою білків-переносників. Іони Н+, що надійшли до клітини, відкачуються протонними насосами на зовнішній бік, а на їх місце надходять іони К+. Потім здійснюється спільний транспорт сахарози та іонів калію по плазмодесмах у порожнини ситоподібних трубок. Аналогічний механізм забезпечує надходження до флоеми амінокислот та інших продуктів фотосинтезу.
Наявність у ситоподібних клітинах іонів калію і таких осмотично активних речовин як сахароза, зумовлює приплив до них води з оточуючих тканин, і у першу чергу – із судин ксилеми. Можливість попадання води разом з іонами К+, Na+, Mg2+ та ін. із судин ксилеми до ситоподібних трубок флоеми забезпечується завдяки наявності між цими провідними тканинами перехідних клітин.
Згідно гіпотези Мюнха, речовини по ситовидних трубках пересуваються у вигляді потоку під тиском від листка до кореня внаслідок осмотичного градієнта між фотосинтезуючими клітинами листка й атрагуючими тканинами. Тиск підтримується у ситоподібних трубках вибірковою проникністю плазмалеми. Це не єдиний механізм транспорту на далекі відстані.
В орієнтації потоку асимілятів провідна роль належить ростучим органам рослини. У рослині можна виділити кілька атрагуючих зон, головними з яких є верхівкова меристема стебла і кореня, дільниці інтеркалярного росту, плоди і запасаючі паренхімні тканини. Роль цих зон у споживанні асимілятів змінюється у онтогенезі рослинного організму.
Мають місце добові зміни транспорту асимілятів. У ранкові години відплив асимілятів загальмований, що, можливо, пов’язане з їх включенням до структурних компонентів хлоропластів, які у нічні години можуть певною мірою деградувати. Має місце і накопичення певної кількості резервних речовин.
Флоемний транспорт асимілятів залежить не тільки від ендогенних факторів. Серед екзогенних факторів температура займає одне з найважливіших місць, оскільки механізм пересування продуктів фотосинтезу залежить від стану цитоплазми, поверхневих мембран і ферментних систем.
Спрямованість та інтенсивність транспорту можна змінити шляхом видалення органів і частин рослин. У практиці сільського господарства з цією метою застосовують чеканку бавовнику, пасинкування томатів, обрізку плодових дерев та ін.
6.10. Залежність інтенсивності фотосинтезу від світла
Про залежність фотосинтезу від світла дає уявлення класичний графік (рис. 41), запропонований К.А. Тимірязєвим. У цьому графіку на осі абсцис розміщені показники освітлення, прийнятого за одиницю; на осі ординат – показники швидкості фотосинтезу у відносних величинах – долях від максимального значення; позначено компенсаційну точку (КТ), тобто, освітлення, при якому фотосинтез і дихання зрівноважуються.
Початковий і середній ступінь (лінійна частина) кривої не лімітовані темновими реакціями фотосинтезу, і тому при зростанні освітлення швидкість процесу збільшується пропорційно. Високий ступінь кривої, навпаки, лімітований темновими реакціями. Тому при ще більшому зростанні освітлення підвищення швидкості фотосинтезу незначне, а у деяких випадках цього збільшення взагалі немає (крива виходить на плато).

Рис. 41. Залежність фотосинтезу від світла (за Тімірязєвим К.А.).
Низка екологічних груп рослин мають особливості світлових кривих. У С4-рослин (кукурудза, цукрова тростина, сорго та ін.), поряд з високою стійкістю до теплового і водного стресу, має місце висока стійкість до інтенсивного освітлення. У тіньовитривалих рослин світлове насичення спостерігається при значно слабшому освітленні, ніж у світлолюбних рослин, компенсаційна точка у них виникає також раніше.
У світлолюбних рослин світлове насичення спостерігається при 60–90 тис. люкс. У тіневитривалих – при значно меншій енергії світла. Наприклад у папоротей воно спостерігається при 5 тисячах люкс. Графічно зв'язок інтенсивність фотосинтезу з інтенсивністю світла можна виразити такими кривими (рис.42).
За оптимальних умов освітлення органічної речовини при фотосинтезі утворюється більше, ніж витрачається на дихання. При недостатньому освітленні може настати момент, коли інтенсивність синтезу органічної речовини врівноважується з інтенсивністю розпаду її на дихання. Тоді спостерігається компенсаційна точка (КТ). Під компенсаційною точкою розуміють інтенсивність світла, при якій інтенсивність відновлення вуглекислого газу під час фотосинтезу врівноважується з інтенсивністю виділення цього газу внаслідок дихання одними і тими ж рослинами за однакових умов.
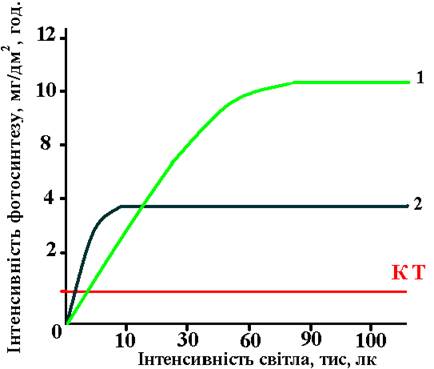
Рис. 42. Залежність фотосинтезу від інтенсивності світла
Дата добавления: 2014-12-26; просмотров: 2995;