Верхній епідерміс; 2 –стовпчаста паренхіма; 3 – губчаста паренхіма; 4 – повітряний простір; 5 – замикаюча клітина; 6 – продих; 7 – ксилема провідного пучка; 8 – флоема.
Должностные обязанности. Принимает грузы со складов в соответствии с сопроводительными документами. Проверяет целостность упаковки (тары). Контролирует наличие приспособлений для перевозки грузов и санитарное состояние соответствующих видов транспорта, предназначенных для перевозки, правильность проведения погрузочно-разгрузочных работ, размещение и укладку грузов. Сопровождает грузы к месту назначения, обеспечивает необходимый режим хранения и сохранность их при транспортировке. Сдает доставленный груз, оформляет приемосдаточную документацию. При необходимости участвует в составлении актов на недостачу, порчу грузов и других документов.
Должен знать: организацию погрузочно-разгрузочных работ; порядок приема и сдачи грузов; адреса основных поставщиков грузов и их складов; условия перевозки и хранения экспедируемых грузов; нормативы простоя подвижного состава и контейнеров под погрузочно-разгрузочными операциями; маршруты перевозки грузов; формы документов на прием и отправку грузов и правила их оформления; основы организации труда; основы законодательства о труде; правила внутреннего трудового распорядка; правила и нормы охраны труда и пожарной безопасности.
Квалификационные требования. Общее среднее образование и специальная подготовка по установленной программе.
верхній епідерміс; 2 –стовпчаста паренхіма; 3 – губчаста паренхіма; 4 – повітряний простір; 5 – замикаюча клітина; 6 – продих; 7 – ксилема провідного пучка; 8 – флоема.
Палісадні клітини містять більшу частину загальної кількості хлоропластів і роблять основний внесок у процес асиміляції вуглекислого газу, який потрапляє у фотосинтезуючі клітини через продихи шляхом дифузії за градієнтом концентрації. Після проходження продихів вуглекислий газ поширюється по системі міжклітинників і надходить у цитоплазму клітин мезофілу у формі вуглецевої кислоти, що утворилася внаслідок сполучення СО2 з водою, яка міститься у пектиновому матриксі клітинних стінок.
Частина вуглекислоти дифундує у хлоропласти через мембрани клітин і хлоропластів, а друга – нейтралізується катіонами цитоплазми і створює відповідний резерв вуглекислого газу у формі бікарбонатів.
6.4.2. Фотосинтетичні пігменти, їх хімічні й фізичні властивості
Вивчення фізико-хімічних властивостей пігментів розпочалося у ХІХст. Ж. Пельтьє і Ж. Каванту у 1818р. отримали з листків спиртовий розчин рослинних пігментів і назвали його хлорофілом.
М.С. Цвєт (1901–1903 рр.), використавши хроматографічний метод аналізу, розділив суміш пігментів на хлорофіл а, в і каротиноїди. У рослині можуть бути фотосинтетичні пігменти трьох груп: хлорофіли, фікобіліни, каротиноїди. У більшості рослин хлорофіл існує у двох формах: синьо-зелений хлорофіл а (С55Н72О5N4Mg) і жовто-зелений в – (С55Н70О6N4Mg). Співвідношення хлорофілів а і в у рослині становить приблизно 3:1, а загальний вміст коливається у межах 0,008–0,8% свіжої речовини. У водоростей, крім хлорофілів а і в, є форми хлорофілів с і d.
Основою структури молекули хлорофілу є магнієвий комплекс порфіринового ядра, у якому атоми азоту чотирьох пірольних кілець пов’язані з атомом магнію, який розміщується у центрі молекули (рис. 33).
Пірольні кільця зв’язані між собою метиновими місточками. До четвертого пірольного кільця приєднаний високомолекулярний спирт фітол – С20Н39ОН, який зумовлює гідрофобні властивості хлорофілу і таким чином – його здатність розміщуватися у ліпідному шарі мембран хлоропластів. Наявні у молекулі хлорофілу десять пар подвійних зв’язків надають їй певної оптичної властивості й високу фотохімічну активність. Завдяки цим зв’язкам і атому магнію хлорофіл має зелений колір.
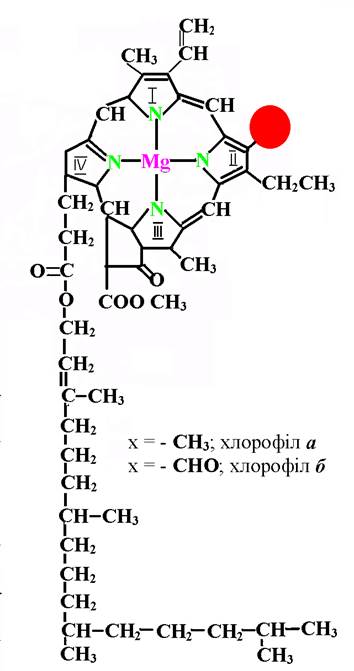
Рис. 33. Структура молекули хлорофілу.
Спектри поглинання хлорофілів а і в мають два чіткі максимуми: у червоній частині спектра – 660 і 640 нм, синьо-фіолетовій – 430–450 нм. Хлорофіл а поглинає енергії синіх променів приблизно у 1,3 рази більше, ніж червоних, а хлорофіл в – у 3 рази. Збуджені під впливом світла молекули хлорофілів після припинення його дії повертаються до початкового стану. Цей перехід супроводжується втратою енергії у вигляді випромінення світла – флуоресценції. Здатність хлорофілу до флуоресценції – одна з найважливіших його властивостей. Вона інтенсивна у розчині хлорофілу і послаблена у тканинах листка. Спиртовий розчин хлорофілу флуоресціює темно-червоним кольором. Послаблення флуоресценції хлорофілу у живих тканинах пов’язане з поглинанням світла від флуоресценції іншими молекулами пігментів.
У рослинних клітинах хлорофіл міститься у хлоропластах, усі структурні компоненти яких складають єдину замкнуту енергетичну систему. Тут постійно відбувається розпад старих і синтез нових молекул хлорофілу. Швидкість оновлення хлорофілу а приблизно у три рази вища, ніж хлорофілу в. Утворення і розпад хлорофілу у живій клітині пов’язані процесами обміну речовин. Головною умовою утворення хлорофілу є наявність світла і хлоропластів, здатних до зеленіння. Хлоропласт формується, росте, старіє і відмирає. Вікові зміни хлоропластів супроводжуються зміною пігментних систем. При старінні листків і, відповідно, хлоропластів, хлорофіл руйнується, листки набувають жовтого забарвлення від наявних каротиноїдів.
Процес оновлення хлорофілу у живих листках зумовлений спадковим, генетичними факторами та умовами навколишнього середовища.
До пігментів, що також беруть участь у фотосинтезі, належать окремі каротиноїди і фікобіліни.
У рослинах зустрічається понад 300 каротиноїдів, але у фотосинтезі беруть участь лише деякі з них. Жовті пігменти є обов’язковими супутниками хлорофілів. У хлоропластах каротиноїдів у три рази менше, ніж хлорофілів. Найважливішими з них є каротин і ксантофіли.
Основною складовою частиною молекул каротиноїдів є вісім сполучених між собою у ланцюг залишків ізопрену:

Елементарна формула каротиноїдів – С40Н56.
Ксантофіли – це каротиноїди, що містять кисень. У молекулах каротиноїдів велика кількість сполучених подвійних зв’язків чергується з одинарними. Таке розміщення зв’язків зумовлює жовтий колір пігментів. За місцем подвійних зв’язків може легко приєднуватися кисень. Каротиноїди поглинають світло у синьо-фіолетовій частині спектра.
До безкисневих каротиноїдів належать супутники хлорофілу a-каротин; b-каротин; g-каротин та ін. До окиснених належать ксантофіли: лютеїн С40Н56О2 – постійний супутник каротину, криптоксантин, міститься у жовтих зернах кукурудзи, шкірці мандарина, зародках пшениці та ін.; рубіксантин С40Н56О (у плодах шипшини), фукоксантин С40Н56О6 (у бурих водоростях); ауроксантінС40Н56О4 (у червоному стручковому перці).
Каротиноїди відіграють роль допоміжних пігментів. Вони передають енергію поглинутих квантів молекулам хлорофілу і цим сприяють більш повному використанню енергії видимої частини спектра світла, яку не поглинає хлорофіл. Крім того, каротиноїди, поглинаючи світло у ділянці високих енергій, виконують захисну функцію і є своєрідним буфером, що запобігає фотоокисленню хлорофілу та інших активних біологічних сполук клітин.
Фікобіліни – це пігменти червоних і синьо-зелених водоростей. До них належать фікоціан С34Н42N4О9 і фікоерітрин С34Н47N4О8. Обидва ці пігменти супутні хлорофілу, але вміст їх значно менший. У хімічному значенні фікобіліни – це комплекс білків і рослинних жовчних кислот. Як допоміжні пігменти фікобіліни виконують роль світлозбиральної антени, забезпечуючи активну передачу поглинутої енергії фотохімічно активним молекулам хлорофілу а.
Фікобіліни поглинають зелену і жовту частину спектра, тобто частину, що знаходиться між двома максимумами поглинання хлорофілу.
При вивченні динаміки фотосинтезу у дослідах з перериванням світла встановлено, що одні ланки реакцій фотосинтезу відбуваються при прямій участі світла, іншим безпосередня участь світла не потрібна. Цим було доведено існування двох фаз фотосинтезу – світлової і темнової.
6.5. Первинні процеси фотосинтезу. Світлова стадія
Система первинних процесів світлової фази фотосинтезу складається з чотирьох послідовних етапів: 1) поглинання квантів світла молекулою пігменту (хлорофілу або іншого) та її збудження; 2) енергія збудженого електрона молекули пігменту розсіюється при флуоресценції або мігрує до іншої молекули хлорофілу (наприклад, реакційного центру) і збуджує її; 3) збуджена молекула передає один з електронів на первинний акцептор і переходить у окислений стан; 4) переданий електрон рухається по електронтранспортному ланцюзі, віддаючи енергію для синтезу АТР або іншому кінцевому акцептору електрона і протона – НАДР+.
Для розуміння механізмів поглинання кванта світла молекулою пігменту і наступної трансформації енергії необхідне знайомство з організацією пігментних систем.
6.5.1. Організація і функціонування 1 та 2 пігментних систем.
У вищих рослин усі фотосинтетичні пігменти містяться у хлоропластах – складних пігменто-білково-ліпоїдних внутрішньоклітинних утвореннях, де безпосередньо відбувається поглинання сонячного світла і його трансформація у органічні сполуки.
У вищих рослин хлоропласти мають еліпсоподібну форму з діаметром 4–6 нм. і товщиною 2–5 нм. Розмір цей непостійний і значно змінюється навіть у однієї і тієї ж рослини. У нижчих рослин хлоропласти більш різноманітні за величиною і формою. Кількість їх дуже варіює, залежно від виду рослин і тканин (від кількох десятків до сотень хлоропластів на одну клітину). Вони мають мембранну будову: зверху покриті подвійною мембраною, з’єднаною з внутрішніми мембранами двох типів: тилакоїдами гран і тилакоїдами строми. Між тилакоїдами міститься колоїдний розчин – строма хлоропласта (рис. 34).
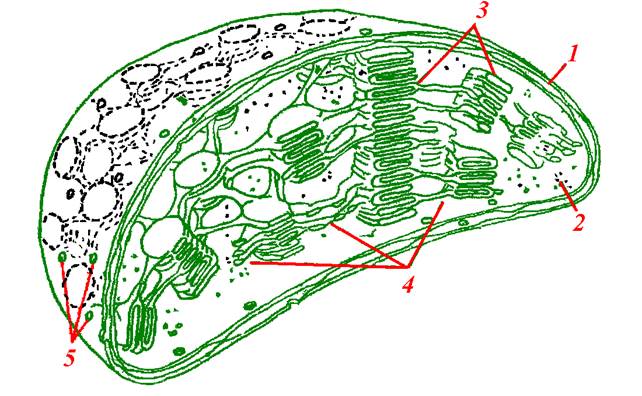
Рис. 34. Будова хлоропласта мезофілу кукурудзи
(модель за Силаєвою А.М.): 1– подвійна мембрана; 2 – строма;
3 – тилаколоїди грани; 4 – тилаколоїди строми; 5 – пори.
На мембранах тилакоїдів розміщені світлозбиральні пігмент-білкові комплекси, електронтранспортні ланцюги фотосистеми 1 (ФС 1) і фотосистеми 2 (ФС 2), АТР-синтетаза і деякі інші ферменти. Ферменти циклу Кальвіна зосереджені у стромі хлоропластів.
Дослідження останніх років свідчать, що більша кількість молекул хлорофілу у хлоропластах міститься у вигляді світлозбиральних антен, а менша – у фотосинтетичних реакційних центрах. Крім хлорофілів а і в, у світлозбиральних антенах містяться каротиноїди. Такий комплекс молекул, поглинаючи енергію світлових квантів, передає її фотохімічно активним молекулам хлорофілу реакційного центру, які набувають при цьому здатності переносити електрони від молекули-донора до молекули-акцептора. Цей процес сполучений з ланцюгом ферментативних реакцій, що призводять до утворення відновлених нікотинамідаденіндинуклеотид-фосфатів (НАДР.Н2) і аденозинтри-фосфатів (АТР).
Відповідно до сучасних уявлень, світлова енергія, необхідна для фотосинтезу, надходить до двох ділянок фотосинтетичного ланцюга переносу електронів за допомогою двох послідовних фотохімічних реакцій. Ділянки ці розміщені у тилаколоїдних мембранах хлоропластів і називаються фотосистемами 1 і 2 (ФС-1, ФС-2). Кожна з них характеризується певним складом молекул хлорофілу і пов’язаних з ними переносників електронів. Енергія квантів світла, поглинутого пігментами світлозбиральних антен, мігрує до реакційних центрів, основою яких є Р700 (фотосистема 1) і Р680 (фотосистема 2). У цих центрах здійснюються реакції фотосинтетичного фосфорилювання – перетворення світлової енергії у лабільну хімічну енергію високоенергетичних сполук АТР і НАДР.Н2.
6.5.2. Електронтранспортні ланцюги фотосистем
Світлозбиральні пігменти-антени, реакційний центр (РЦ) і відповідні ферменти електронтранспортного ланцюга утворюють комплекс – фотосинтетичну одиницю. У вищих рослин фотосинтетична одиниця має 250–300 молекул хлорофілу і 500 молекул каротину. Відомо кілька моделей організації фотосинтетичної одиниці (ФСО). Окремі пігментні білкові комплекси фотосистем можуть працювати незалежно одна від одної. При цьому кожний реакційний центр може отримувати енергію тільки від своїх світлозбиральних молекул. Це моно- або уніцентральна модель. Інколи окремі світлозбиральні комплекси, що обслуговують реакційний центр, можуть утворювати домени, між якими можливе перенесення енергії. Це буде мультицентральна фотосинтетична одиниця, різні реакційні центри якої пов’язані між собою через міграцію енергії. При цьому у кожному домені може бути 20–30 РЦ. Фотосинтетична одиниця не завжди є однорідною структурою. Комплекс ФС-1, наприклад, включає 5–7 поліпептидів і 100–200 молекул хлорофілу а на один РЦ Р700. Ці молекули створюють ядро ФС-1 – 40 молекул хлорофілу; внутрішню антену – 20 молекул і периферійну антену – 40 молекул хлорофілу.
Реакційний центр (РЦ) складається з білка, молекул хлорофілу, феофітину, кількох цитохромів, фередоксину і НАД. Для функціонування РЦ важливу роль має наявність у ньому агрегатів хлорофілу і феофітину, в яких електрон від збудженої молекули хлорофілу передається до збудженої молекули феофітину.
При нормальних фізіологічних умовах розміщення у хлоропластах білково-пігментних комплексів є нерівномірним: ФС-2 локалізується у зонах щільного стикування тилакоїдів гран; ФС-1 знаходиться переважно у тилакоїдах строми. Зв’язок між просторово віддаленими комплексами здійснюється за допомогою рухомих переносників електронів –пластохінону, пластоціаніну і фередоксину, які можуть переміщуватися шляхом дифузії, і забезпечувати перенесення електронів між ФС-1 і ФС-2.
Поглинання квантів світла відбувається послідовно кожною молекулою світлозбиральної антени зі швидкістю один квант за 0,1 с. Енергія поглинутих фотонів світла стікається до РЦ.
Пропускна спроможність фотосинтетичної одиниці (світлове насичення фотосинтезу) визначається здатністю реакційного центру поглинати 50 квантів світла за 1 секунду.
Донором електронів для ФС-1 є хлорофіл реакційних центрів Р700 і цитохром, а для ФС-2 – вода і хлорофіл реакційних центрів Р680.
Природа первинних акцепторів електронів ФС-1 остаточно не вивчена. Вважається, що ними можуть бути залізо-сірчані білки. Нечіткою є точка зору і про природу акцепторів електронів ФС-2. Припускається, що акцепторами останніх є феофітин. На окислений РЦ ФС-2 електрони надходять від води через спеціальну систему фотолізу води і виділення кисню.
Переносники електронів локалізуються у хлоропластах.
Від збудженого хлорофілу Р700 електрони надходять до первинних акцепторів ФС-1, потім до фередоксину. У процесі циклічного транспорту електрони від фередоксину переносяться на цитохром b6 і потім – на пластохінон. Далі, як і при нециклічному транспорті, електрони переносяться на залізо-сірчаний центр, цитохром f, пластоціанін і Р700.
Найбільш повільною стадією цих процесів є стадія окислення пластохінону, яка дозволяє змінювати загальну швидкість транспорту електронів зміною швидкості окислення пластохінону, що є можливим при нестачі субстратів фотофосфорилювання. Пластохінон відіграє основну роль у перенесенні протона із зовнішнього до внутрішнього простору тилакоїдів при освітленні хлоропластів.
6.5.3. Механізм участі у процесі перетворення сонячної енергії
Суть світлової стадії фотосинтезу полягає у перетворенні енергії квантів світла в енергію хімічнх лабільних високореактивних сполук – АТР і НАДР.Н2. Утворення їх відбувається послідовно під час фотофізичного і фотохімічного етепів.
Ефективність фотофізичного етапу визначається енергією кванта світла, поглинутого молекулою хлорофілу реакційного центру. При поглинанні світла молекула переходить у збуджений стан, тобто один з її електронів потрапляє на більш високий енергетичний рівень. Якщо поглинуто квант синього світла (65 кДж.), то електрон у молекулі хлорофілу з основного (S0 рівня) підніметься до синглетного рівня – S2* (рис. 35).
Такий стан енергетично нестійкий, і електрон майже у ту ж мить (через 10-12 – 10-9 сек) повертається до основного (початкового) рівня. Енергія збудження на зворотному шляху від S2* до S1* частково перетворюється на тепло, а на ділянці S1* – S0 розсіюється у вигляді тепла або флуоресценції і практично не перетворюється в енергію хімічних зв’язків.
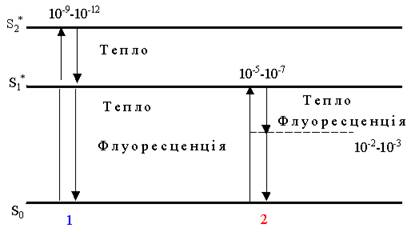
Рис. 35. Рівні збудження хлорофілу під впливом світла різної довжини хвилі:
1– поглинання кванта синього світла; 2 – поглинання кванта червоного світла;
При поглинанні кванта червоного світла (44 кДж) електрон з S0-рівня потрапляє на більш низький, ніж при синьому світлі S1*-рівень (рис. 34), знаходиться на ньому дуже короткий відрізок часу (10-5 – 10-7 сек) і повертається на початковий рівень S0, віддаючи енергію збудження у вигляді тепла або флуоресценції. На зворотному шляху спин його може змінитися на протилежний, тобто стати таким, як і в електрона, що знаходиться на S0-рівні. Згідно із законом заборони Паулі, на одній орбіталі не можуть одночасно знаходитися два електрони з однаковим спином. Тому електрон, який повертається з S1* до S0, затримується на триплетному рівні Т до того часу, доки знак його спину не відновиться до початкового. На це витрачається 102 – 103 сек. Цього часу цілком достатньо, щоб електрон, який знаходиться на триплетному рівні, встиг прореагувати з молекулою хімічного акцептора - відповідного ферменту і передати йому частину енергії, яка буде витрачена на утворення певної енергетичної сполуки – АТР і НАДР.Н2 . Якщо цього не трапиться, то енергія збудження теж розсіюється у вигляді тепла або флуоресценції.
Таким чином, фотофізичний етап – це етап, на якому енергія світлового випромінення перетворюється в енергію збудженого електрона молекули хлорофілу реакційного центра.
У подальших фотохімічних реакціях беруть участь тільки електрони молекул хлорофілу, що знаходяться у довгоживучому (триплетному) стані збудження. На фотохімічному етапі енергія збудженого електрона перетворюється у хімічно пов’язану енергію сполук АТР і НАДР.Н2 . Процес утворення цих сполук відбувається у такій послідовності:
а) фотоліз води – розщеплення води, індуковане хлорофілом, окисленим під впливом світла: 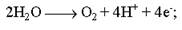
б) фотосинтетичне фосфорилювання – циклічне і нециклічне.
6.5.4. Фотоліз води
Фотоліз води – це реакція розщеплення води, яка спостерігається при освітленні хлоропластів. У її здійсненні безпосередню участь бере хлорофіл. Фотохімічне розщеплення води було вперше виявлене Р.Хіллом у 1937р. При освітленні суспензії хлоропластів у присутності акцептора електронів (А)
виділявся кисень: 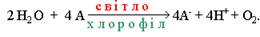
На честь ученого, що відкрив це явище, воно було назване реакцією Хілла.
Природнім акцептором електронів у цій реакції є нікотинамідаде-ніндинуклеотидфосфат (НАДР+).
Та обставина, що реакція Хілла здійснюється без участі СО2, свідчить, про те, що кисень, який виділяється при фотосинтезі, є киснем води, а не вуглекислого газу, як це вважалося раніше.
Фотоліз води здійснюється шляхом відщеплення електронів (фотоокислення) від молекул води з наступним виділенням протонів і вільного молекулярного кисню:
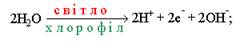


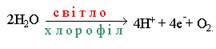
Електрони і протони, що утворилися у реакції Хілла, використовуються у світловій стадії фотосинтезу для відновлення НАДР+ до НАДР.Н2 і заповнення електронної дірки у ФС-1 при нециклічному фотосинтетичному фосфорилюванні.
6.5.5. Фотосинтетичне фосфорилювання
Фотосинтетичне фосфорилювання – це процес утворення високоенергетичних сполук шляхом трансформації енергії електрона. Здійснюється воно за участю двох фотосистем – ФС-1 і ФС-2. Рух збуджених електронів відбувається по відповідному електронтранспортному ланцюзі (рис. 36).
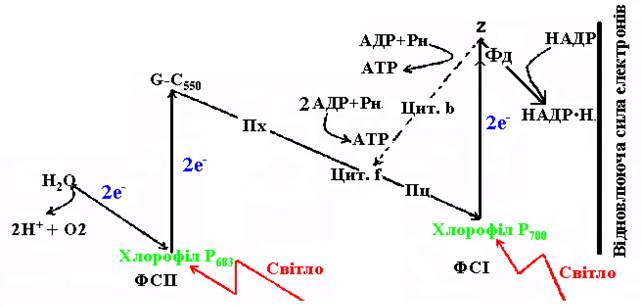
Рис. 36. Схема руху електронів при світловій стадії фотосинтезу.
Розрізнюють два види фотосинтетичного фосфорилювання – циклічне і нециклічне.
Циклічне фотосинтетичного фосфорилювання – це процес утворення АТР при переміщенні електрона по замкнутому колу ЕТЛ. Воно здійснюється за участю фотосистеми 1 (ФС-1), реакційним центром якої є молекули хлорофілу а з максимальним поглинанням світла з довжиною хвилі 700 нм. (Р700). При попаданні кванта світла на молекулу хлорофілу реакційного центра електрон, що вибивається з орбіталі енергією цього світла, проходить шлях по ЕТЛ у напрямі зростання окислювального потенціалу і на певному відрізку шляху вивільнює енергію, яка використовується для синтезу двох молекул АТР. Сам електрон продовжує шлях, повертається на попереднє місце у своїй орбіталі й заповнює там електронну дірку, що утворилася при його вибиванні квантом світла. Після цього хлорофіл реакційного центру може знову поглинати фотони і переходити до збудженого стану – цикл повторюється.
При неціклічному фотосинтетичному фосфорилюванні працюють обидві фотосистеми. У фотосистемі-2 реакційним центром є хлорофіл з максимумом поглинання світла з довжиною хвилі 683 нм. (Р683).
Поглинаючи квант світла, молекула реакційного центру ФС-2 вивільнює свій електрон, і в ній утворюється електронна дірка. Електронний ланцюг, який поєднує ФС-1 і ФС-2, розпочинається з акцептора електронів – ферменту Q ФС-2. Послідовність руху елетронів між ФС-2 і ФС-1 така: акцептор (Q) ® ¾® пластохінон (Пх) ¾® цитохром ¾® пластоціанін (Пц) ¾®Р700. Проте, потрапити до хлорофілу Р700 електрон може лише після того, як Р700, прийнявши квант світла, пошле по ланцюзі свій електрон. Це призведе до утворення електронної дірки, яка і буде заповнена електроном, що рухається від Р683. Елетрон від молекули хлорофілу Р700 рухається до переносника Z, передається далі на більш окисленні сполуки – фередоксин (Фд) і виводиться на зовнішній бік тилакоїду хлоропласта. Тут він пов’язується з протонами і відновлює НАДР до НАДР.Н2. Постачальником протонів є вода, що піддається фотолізу. Вона також є донором електронів для заповнення електронної дірки у Р683. Отже, молекула хлорофілу ФС-2 повертається до вихідного стану за рахунок електронів, які утворилися при фотолізі води.
Під час нециклічного фосфорилювання утворюються два види сполук - АТР і НАДР×Н2. Перша – при русі електрона від Р683 до Р700, а друга – від Р700 до зовнішнього боку тилакоїдів. У зв’язку з тим, що при нециклічному фосфорилюванні працюють одночасно обидві фотосистеми, за один прохід електронів утворюються чотири молекули АТР і дві молекули НАДР.Н2. АТР слугує джерелом енергії, НАДР.Н2 – джерелом енергії і водню (відновна сила) для відновлення СО2 до вуглеводів у реакціях темнової стадії фотосинтезу.
Механізм утворення АТР найбільш вдало пояснюється хеміосмотичною теорією Мітчелла (1961-1966 рр.), згідно з якою на одному боці фотосинтетичної мембрани накопичуються протони, створюючи різницю потенціалів, і, як наслідок – електричне поле, що має відповідний потенціал енергії. У результаті періодичного підключення до цього процесу ферменто АТР-ази концентрація протонів з обох боків мембрани вирівнюється, а енергія, яка при цьому вивільнюється, витрачається на утворення АТР з АДР і неорганічного фосфору (Рн). Ця теорія має назву хеміосмотичної теорії сполучення, тому що осмотична енергія, нагромаджена у вигляді різниці електрохімічних потенціалів протонів Н+, витрачається на хімічну роботу – синтез АТР.
6.6. Темнова стадія. Метаболізм вуглецю при фотосинтезі
Відновлення вуглецю до вуглеводів відбувається за допомогою енергії АТР і “відновлювальної сили” – НАДР*Н2, без прямої участі сонячного світла і має назву темнової стадії фотосинтезу.
6.6.1. Цикл Кальвіна. С3 – шлях фотосинтезу
Ця стадія фотосинтезу була детально вивчена у 1946–1956рр. американським біохіміком М. Кальвіном з співробітниками яка відома також під назвою циклу Кальвіна.
Прослідкувати весь цикл реакцій темнової стадії фотосинтезу стало можливим завдяки використанню у наукових дослідженнях радіоактивного вуглецю у поєднанні з паперовою хроматографією.
Слід нагадати, що принцип хроматографії розробив і вперше використав у біохімічних дослідженнях російський учений М.С. Цвєт.
У дослідах американських учених з міченою вуглекислотою вже через 2 с після введення цієї сполуки у суспензію водорості хлорели або хлоропластів, радіоактивний вуглець виявили у 3-фосфогліцериновій кислоті (ФГК). Дещо пізніше (через 7 с) радіоактивна мітка локалізувалася у монофосфатах і дифосфатах цукрів. Це дало підставу стверджувати, що первинними продуктами фотосинтезу є фосфогліцеринова кислота, а акцептором СО2 – рибулозо-1,5-біфосфат (РБФ).
Приєднання вуглекислоти до акцептора і подальші перетворення її до цукрів мають складний характер, вимагають участі енергії і певних ферментів і відбуваються у три послідовні етапи: 1) карбоксилювання акцептора; 2) відновлення вуглекислого газу; 3) регенерація рибулозобіфосфату.
Перший етап полягає у приєднанні СО2 до акцептора за участю ферменту рибулозо-1,5-біфосфаткарбоксилази. Внаслідок цього утворюється нестійка шестивуглецева сполука, яка миттєво гідролізується до двох молекул фосфогліцеринової кислоти (ФГК):
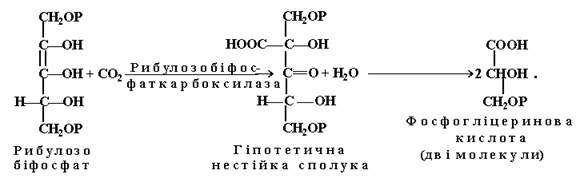
На другому етапі темнової стадії за участю енергії АТР і “відновлювальної сили” - НАДР×Н2 утворюється глюкоза, а саме: до кожної з двох молекул ФГК за допомогою ферменту фосфогліцеринкінази приєднується по одному залишку ортофосфорної кислоти, внаслідок чого утворюється дифосфогліцеринова кислота, яка під впливом ферменту дегідрогенази і НАДР.Н2 відновлюється до фосфогліцеринового альдегіду (ФГА). Одна з молекул цієї сполуки при участі тріозофосфат-ізомерази ізомеризується до диоксиацетонфосфату (ДОАФ). Із двох тріоз - ФГА і ДОАФ – за допомогою альдолази здійснюється синтез однієї молекули шестивуглецевої сполуки – фруктозо-1,6-біфосфату (ФБФ), яка є вихідною сполукою для утворення інших вуглеводів (сахарози, крохмалю):
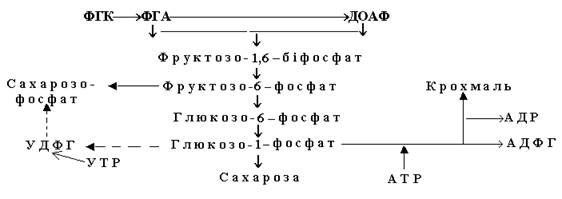
Заключний, третій етап циклу Кальвіна полягає у регенерації рибулозо-1,5-біфосфату. Він складається з ряду реакцій, у яких з 3,4 і 7-ми вуглецевих сполук утворюються пентози. У одній із перших реакцій від фруктозо-6-фосфату під дією транскетолази відокремлюються два атоми вуглецю і утворюється еритрозо-4-фосфат. Ця сполука конденсується з ДОАФ за участю альдолази і синтезується цукор - седогептулозобіфосфат (СДФ). Від нього відокремлюється ортофосфат, внаслідок чого утворюється седогептулозо-7-фосфат, який набуває транскетолазної активності з відокремленням двох атомів вуглецю. Виникає нова сполука – пентоза рибозо-5-фосфат. За допомогою ізомерази ця сполука перетворюється у рибулозо-5-фосфат.
Фонд пентоз поповнюється також за рахунок транскетолазних реакцій: двовуглецевий компонент приєднується до фосфогліцеринового альдегіду і утворюється пентоза ксилулозо-5-фосфат. Завершальним процесом етапу є перетворення пентоз до рибулозобіфосфату. Він утворюється з фосфомонопентоз, які повторно фосфорилюються за рахунок АТР, що надходить від світлової стадії фотосинтезу.
Із 12 молекул ФГА на синтез вуглеводів використовуються дві, а основна їх кількість (10 молекул) витрачається на відновлення акцептора СО2, тобто рибулозобіфосфату.
Завдяки цьому рослина досить швидко нагромаджує велику кількість молекул акцептора і тим посилює інтенсивність поглинання СО2.
Регенерований рибулозо-1,5-фосфат приєднує молекулу СО2 і знову підключається до циклу.
У циклі Кальвіна відбувається перетворення тріоз, тому рослини з таким циклом фотосинтезу називаються С3-рослинами, а шлях фотосинтезу – С3-шляхом фотосинтезу. Переважна більшість рослин (близько 80%) мають такий шлях фотосинтезу. Спрощена схема циклу Кальвіна показана на рис. 37.
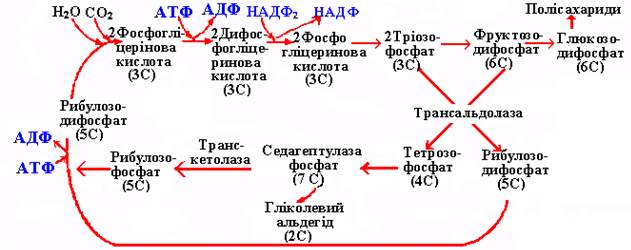
Рис. 37. Схема С3-шляху фотосинтезу (Лібберт Е, 1976, з деякими змінами).
6.6.2. Цикл Хетча-Слека. С4-шлях фотосинтезу
Крім фіксації СО2 у пентозофосфатному циклі (цикл Кальвіна), процес карбоксилювання здійснюється і при взаємодії вуглекислоти з монокарбоновими кислотами шляхом утворення дикарбонових кислот.
При вивченні кінетики і продуктів фотосинтезу у рослин тропічного походження (кукурудза, сорго, цукрова тростина) і родини товстолистих виявлено інший тип фіксації вуглекислого газу.
Встановлено, що у хлоропластах клітин обкладки провідних пучків у першу секунду освітлення фіксація і перетворення СО2 здійснюються за циклом Кальвіна з утворенням тріоз, а у хлоропластах мезофільної тканини первинними продуктами фотосинтезу є чотиривуглецеві сполуки – малат і аспартат.
Цей тип фотосинтезу вперше був вивчений австралійськими вченими М.Д.Хетчем і К.Р.Слеком і отримав назву циклу Хетча-Слека. Рослини з таким типом фотосинтезу називаються С4-рослинами, а шлях вуглецю при фотосинтезі – С4-шляхом.
С4-рослини мають низку особливостей в анатомічній будові і функціях тканин листків та фотосинтетичного апарату. Листкова пластинка у них густо пронизана сіткою провідних судинних пучків, оточених клітинами обкладкової паренхіми з великою кількістю крупних хлоропластів. Клітини ж мезофілу мають хлоропласти звичайного виду.
У хлоропластах клітин обкладкової тканини функціонує цикл Кальвіна, а у хлоропластах мезофілу – цикл Хетча-Слека. Цей цикл включає такі послідовні процеси:
а) карбоксилювання акцептора – фосфоенолпіровиноградної кислоти (ФЕП) з утворенням щавлевооцтової кислоти (ЩОК);
б) відновлення ЩОК до яблучної кислоти (малат);
в) декарбоксилювання малату до піровиноградної кислоти (ПВК);
г) новоутворення ФЕП з ПВК за допомогою енергії АТР.
Вуглекислий газ, що потрапляє до клітин мезофілу, вступає у реакцію з ФЕП за участю ферменту фосфоенолпіруваткарбоксилази. Наслідком реакції є чотиривуглецева сполука – щавлевооцтова кислота (ЩОК). Ця сполука за рахунок НАДР*Н2 відновлюється до яблучної кислоти, яка переноситься до хлоропластів клітин обкладки судинного пучка. Це можливе завдяки інтенсивному обміну продуктами первинного перетворення вуглекислого газу між хлоропластами клітин мезофілу і обкладок судинного пучка. У хлоропластах клітин обкладок яблучна кислота декарбоксилюється, тобто віддає молекулу СО2, яка вступає у цикл Кальвіна, що функціонує у цих клітинах. Піровиноградна кислота, що утворилася внаслідок декарбоксилювання яблучної кислоти, переміщується по численних плазмодесмах знову до клітин мезофілу, у яких за допомогою енергії АТР перетворюється у ФЕП, тобто у первинний акцептор СО2. Цикл повторюється. Схема реакцій циклу Хечта-Слека (рис. 38).
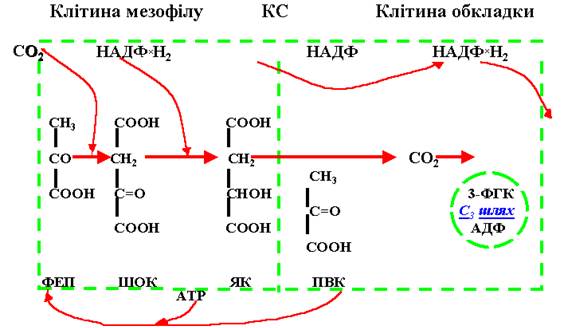
Рис. 38. С4-шлях фотосинтезу у листках кукурудзи. КС – клітинна стінка.
Цикл Хетча-Слека не замінює цикл Кальвіна, а лише доповнює його. Тому у С4-рослин злагоджено працюють обидва цикли. Компартментація процесів асиміляції СО2 дозволяє таким рослинам здійснювати фотосинтез і при закритих продихах, використовуючи вуглекислий газ, що виділяється при декарбоксилюванні яблучної кислоти. С4-рослини, як правило, мають більш високу продуктивність, ніж С3-рослини. Вони можуть здійснювати фотосинтез при більш низьких концентраціях СО2 і при високій інтенсивності світла.
Вважають, що у С4-рослин відсутнє фотодихання, на яке у С3-рослин при високій температурі у сонячні дні витрачається до 50% НАДР.Н2, внаслідок чього близько 50% СО2 у такі дні виділяється в атмосферу і не використовується на синтез органічної речовини. У С4-рослин вуглекислота, що виділяється, у тканинах використовується вдруге.
6.6.3. Інші шляхи перетворення вуглецю при фотосинтезі
Крім розглянутих типів фіксації СО2 і перетворення первинних продуктів існує ще один шлях фотосинтезу – САМ-шлях (Сrassulaceае acid Metabolіsmus). Він уперше був виявлений у рослин родини товстолистих – Сrassulaceae. Це шлях перетворення вуглецю у рослин-сукулентів, що ростуть в умовах різко посушливого клімату і виживають завдяки більш ефективному використанню води, яку поглинають вночі. Для різкого зменшення втрат вологи їх продихи вдень закриті. Тому СО2 може проникати у тканини листка лише вночі і за участю фосфоенолпіруваткарбоксилази взаємодіяти з ФЕП, утворюючи за циклом Хетча-Слека щавлевооцтову кислоту. Джерелом ФЕП є цукри. Вуглекислий газ, утворений при диханні, теж використовується на карбоксилювання ФЕП. Синтезована таким шляхом ЩОК відновлюється до яблучної кислоти (ЯК) і зосереджується у вакуолях клітин листка (рис. 38).
Удень при високій температурі й дефіциті вологи продихи у рослин закриті. Яблучна кислота з вакуолей транспортується до цитоплазми, де і відбувається її декарбоксилювання з утворенням піровиноградної кислоти і СО2. Вуглекислий газ включається у процес фотосинтезу за циклом Кальвіна. Отже, у сукулентів діють обидва типи фіксування СО2 - С3-тип (цикл Кальвіна – удень) і С4-тип (цикл Хетча-Слека – уночі). Але якщо у С4-рослин ці шляхи фотосинтезу мають просторове розмежування (С3-шлях відбувається у хлоропластах обкладки, а С4-шлях – у хлоропластах мезофілу), то при САМ-шляху має місце часове розмежування, тобто вночі здійснюється фотосинтез за циклом Хетча-Слека, а вдень – за циклом Кальвіна.
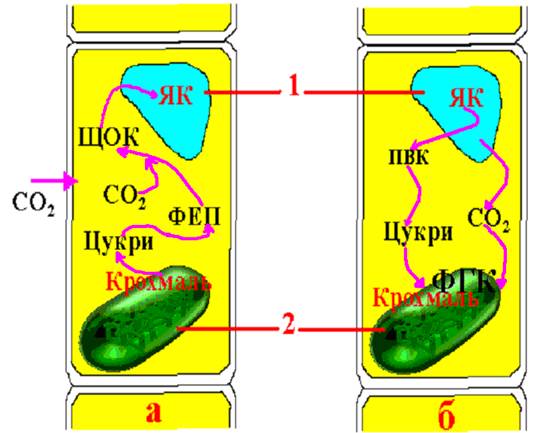
Рис. 38. Метаболізм вуглецю у рослин з САМ-шляхом фотосинтезу:
а – вночі (продихи відкриті); б – вдень (продихи закриті); 1 – вакуолі; 2 – хлоропласти.
Продуктивність фотосинтезу САМ-шляху невисока. Це пов’язано, здебільшого, з несприятливими умовами росту сукулентів.
6.6.4. Фотодихання
Значну частину поглинутої листком енергії С3-рослини витрачають на фотодихання, яке виникає внаслідок активованого світлом окислення киснем рибулозобіфосфату до фосфогліцеринової і гліколевої кислот. Фотодихання супроводжується виділенням СО2. У зв’язку з тим, що первинним продуктом цього процесу є гліколева кислота, то процес фотодихання називається також гліколатним. Синтезу гліколату сприяють і високі концентрації кисню. В окремих С3-рослин з низькою інтенсивністю фотосинтезу інтенсивність фотодихання може досягати 50% від інтенсивності фотосинтезу.
Доведено, що фотодихання основане на реакції, яка каталізується ключовим ферментом циклу Кальвіна – рибулозодифосфаткарбоксилазою (оксигеназою). Кисень і вуглекислий газ конкурують за активний центр ферменту. Тому швидкість реакцій визначається концентраціями цих газів, температурою і значенням рН середовища. За нормальних умов карбоксилазна активність рибулозобіфосфаткарбоксилази у три–п’ять разів вища за оксигеназну.
Фотодихання з високою за СО2 компенсаційною точкою відмічено у великої групи вищих рослин (соняшнику, пшениці, тютюну, бобових та ін.). У рослин з низькою за СО2 компенсаційною точкою фотодихання майже відсутнє (кукурудза, сорго, цукрова тростина та інші; С4-рослини). Воно здійснюється при взаємодії трьох органел–хлоропластів, пероксисом і мітохондрій за рахунок відновлювальної сили НАДР.Н2, утвореної при світловій стадії фотосинтезу.
Основним субстратом фотодихання є гліколева кислота. Вона надходить до пероксисом, де окислюється киснем з утворенням перекису водню і гліоксилевої кислоти:

Перекис водню під впливом каталази у пероксисомі розкладається, а гліоксилева кислота амінується з утворенням амінокислоти гліцин. Частина молекул гліоксилевої кислоти може мігрувати від пероксисом до хлоропластів і відновлюватися знову до гліколевої кислоти. Гліцин з пероксисом транспортується до мітохондрій, де перетворюється у серин з виділенням СО2.
Гліоксилатний шлях у С3-рослин може завершуватися у мітохондріях. Фотодихання знижує чисту продуктивність фотосинтезу.
У С4-рослин утворений при фотодиханні двоокис вуглецю у клітинах мезофілу карбоксилює ФЕП і далі відновлюється до вуглеводів за схемою С4-фотосинтезу.
Роль фотодихання полягає у тому, що при гліколатному шляху відбувається синтез амінокислот гліцину і серину.
6.7. Системи регуляції фотосинтезу
Механізм регуляції фотосинтезу досить складний. На рівні цілого організму можна виділити ендогенний, екзогенний і метаболічний типи регуляції. Ендогенний тип базується на взаємодії джерела і споживача асимілятів. Існує зв’язок між інтенсивністю фотосинтезу листка та експортом з нього продуктів асиміляції. До факторів ендогенної регуляції належать різнобічні зворотні зв’язки між фотосинтезом, як джерелом асимілятів, і ростовими процесами, як споживачами цих асимілятів. Велике значення у ендогенній регуляції належить фітогормонам. Деякі фітогормони синтезуються у хлоропластах і беруть безпосередню участь у регуляції інтенсивності фотосинтезу шляхом впливу на механізм руху продихів, з яким пов’язане надходження СО2 до мезофілу листка. Як відомо, у динаміці руху продихів важливе місце належить абсцизовій кислоті. Гіберелова кислота у ізольованих хлоропластах активує фосфорилювання, а цитокініни ініціюють синтез білка.
Екзогенний тип реакції виявляється при взаємодії між живим організмом і факторами навколишнього середовища, до яких належать температура, волога, газовий склад атмосфери, елементи мінерального живлення та деякі інші.
Дата добавления: 2014-12-26; просмотров: 3897;