у світлолюбних (1) і тіньовитривалих (2) рослин.
КТ – компенсаційна точка
У світлолюбивих рослин КТ спостерігається при 800–2000 люкс, а у тіневитривалих – при 250–300 люкс. При освітленні нижче компенсаційної точки розпад органічної речовини перевищує його синтез і ріст рослини призупиняється.
Це пов’язане з тим, що тіньовитривалі рослини характеризуються невеликою інтенсивністю дихання. За умов слабкого освітлення швидкість фотосинтезу вища у тіньовитривалих рослин і, навпаки, при інтенсивному освітленні – у світлолюбних.
Фотосинтез можливий при мінімальному освітленні. У більшості рослин інтенсивність фотосинтезу зростає при збільшенні освітлення до 1/3 повного сонячного світла, після чого настає світлове насичення, тобто крива фотосинтезу виходить на плато і навіть може спадати. Це пояснюється тим, що при насиченні світлом фотосинтетичного апарату виникає надлишкова кількість збуджених молекул хлорофілу, енергія яких не може бути включеною до фотохімічних реакцій, а використовується у неспецифічних фотореакціях, на фотоокислення та ін.
Зелена рослина має велику пластичність щодо інтенсивності світла.
Крім інтенсивності, велике значення для фотосинтезу має спектральний склад світла. Видиму частину сонячного спектра (380–720 нм) прийнято відносити до фотосинтетично активної радіації (ФАР), яка складається з прямого і розсіяного сонячного світла. Частки ФАР та інфрачервоної радіації у складі сонячного світла приблизно рівні, але їх поглинання і пропускання дуже різні: видимі промені поглинаються на 85 відсотків; інфрачервоні - лише на одну чверть; перші пропускаються тільки на 5 відсотків, другі – на 30. Особливо велика кількість (45%) інфрачервоних променів відбиваються, тоді як видимих – лише 10% (рис. 43).
Листки рослин можуть розміщуватися у просторі так, що використовують як пряме, так і розсіяне світло.
Швидкість фотосинтезу у різних ділянках спектра різна. Найбільш висока швидкість фотосинтезу припадає на довгохвильову, червону частину спектра. Це пов’язано з тим, що енергії одного кванта червоного світла достатньо для переведення електрону молекули хлорофілу на перший синглетний рівень збудження (S1*), що забезпечує можливість наступного використання її у фотофізичних реакціях. Фотони синього світла несуть набагато більше енергії, яка викликає більш високий рівень збудження (S2*). Надлишок енергії при поверненні молекули із збудженого до початкового стану перетворюється у теплову, тобто спостерігається не ефективна витрата енергії.
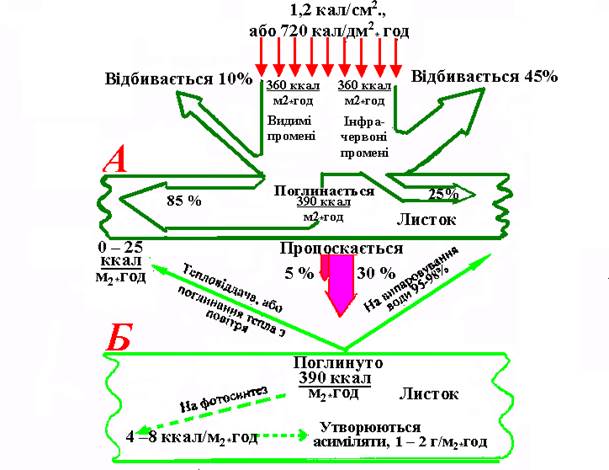
Рис. 43. Енергетичний баланс листка при попаданні на нього сонячної енергії.
А– загальний розподіл енергії; Б – розподіл поглинутої енергії (за Ничипоровичем О.А, 1965).
Червоні промені завжди наявні у сонячному світлі. Кількість їх у прямих сонячних променях змінюється протягом світлового дня. Уранці переважає розсіяне світло, представлене переважно (майже 2/3) довгохвильовими червоними променями. У полуденні години червоні промені становлять не більше 1/4 від загальної кількості сонячного світла. Це є однією з основних причин зниження ефективності фотосинтезу у полуденні години.
Якісний спектральний склад світла впливає не тільки на інтенсивність фотосинтезу, але й на хімічний склад асимілятів. Встановлено, що при синьому світлі у листках, крім вуглеводів, утворюються і невуглеводні продукти (органічні кислоти та ін.), і швидкість фотосинтезу зростає.
6.11. Вплив інших факторів на інтенсивність фотосинтезу
Вуглекислота. Вміст вуглекислоти у атмосфері відносно невеликий, досить постійний і становить близько 0,03 об’ємних відсотки. Безпосередньо ж у посівах різних сільськогосподарських культур у різні періоди доби концентрація СО2 непостійна: вона то знижується внаслідок поглинання рослинами у процесі фотосинтезу, то зростає при зниженні інтенсивності фотосинтезу. Певне значення для динаміки вуглекислоти у надземному шарі повітря має швидкість мінералізації гумусу, органічних решток і органічних добрив.
Зростання концентрації вуглекислоти в атмосфері, навіть десятикратне, сприяє поступовому прискоренню процесу фотосинтезу. Підживлення рослин вуглекислим газом у закритих приміщеннях є досить ефективним заходом для підвищення продуктивності, особливо С3-рослин. С4-рослини не реагують на збагачення повітря вуглекислим газом, тому що у них існує особливий механізм накопичення СО2 у тканинах листків.
Температура. Залежність швидкості видимого фотосинтезу від температури зображується куполоподібною кривою. При цьому пригнічення фотосинтезу розпочинається нерідко при відносно невисоких температурах, після 20 оС (рис. 44).
 Рис. 44. Залежність фотосинтезу від температури листка (за Гейте, Джонсон та ін., 1972): 1 – бавовник; 2 – соняшник; 3 - сорго.
Рис. 44. Залежність фотосинтезу від температури листка (за Гейте, Джонсон та ін., 1972): 1 – бавовник; 2 – соняшник; 3 - сорго.
Крива фотосинтезу має три кардинальні температурні точки: мінімальну, при якій фотосинтез розпочинається; оптимальну – інтенсивність фотосинтезу досягає найвищих показників; максимальну, при якій спостерігається різке падіння і припинення фотосинтезу.
Найголовнішою причиною такої залежності асиміляції СО2 є неоднакова реакція на температурний режим видимого фотосинтезу і дихання, а саме: зі зростанням температури інтенсивність дихання зростає значно швидше, ніж інтенсивність дійсного фотосинтезу. З іншого боку, під впливом нагрівання чутлива структура хлоропластів, у першу чергу біологічних мембран, зазнає денатураційних змін, у тому числі незворотніх.
Температурні показники кардинальних точок проходження фотосинтезу неоднакові у рослин різних грунтово-кліматичних зон: мінімальні – при +5 оС у тропічних і субтропічних; для більшості рослин помірної зони – близько 0 оС; у деяких хвойних (сосна, ялина) – -2, -3 оС. Максимум знаходиться між 35 і 50 оС.
Вплив різних температур на фотосинтез особливо позначається при інтенсивному освітленні. Слід зазначити існування досить складної залежності фотосинтезу від світла, концентрації СО2 і температури. Дуже тісним є взаємозв’язок світла (дія на фотохімічні реакції) і температури (дія на швидкість ензимних реакцій). Швидкість ензимних реакцій у ланцюзі відновлення вуглекислоти при підвищенні температури на 10 оС зростає удвічі. Тому при високій інтенсивності світла низькі температури гальмують ферментативні реакції і є лімітуючими факторами для фотосинтезу. При низькій інтенсивності світла фотосинтез не залежить від температури, оскільки стримується швидкістю фотохімічних реакцій.
Вода відіграє першочергове значення у здійсненні рослиною фотосинтетичних функцій. Це значення визначається передусім тим, що вода, як вихідна сполука, є безпосереднім учасником процесу фотосинтезу.
Умови водного режиму значною мірою визначають структуру хлоропластів, впливають на біосинтез і вміст пігментів у листках рослин, на утворення листкової поверхні.
Тривалий водний дефіцит призводить до порушення нециклічного фотофосфорилювання. Водночас частковий дефіцит води (5–20 відсотків від повного насичення) забезпечує оптимальний хід фотосинтезу.
Мінеральне живлення. Повітряне і кореневе живлення тісно взаємопов’язані. Залежність фотосинтезу від мінеральних елементів визначається їх необхідністю для формування фотосинтетичного апарату (пігментів, фотосинтетичних систем та ін.), його оновлення і функціонування.
Дослідами Національного аграрного унівеситету доведено, що тилакоїди, ламелярно-гранулярна структура хлоропластів мезофілу листків кукурудзи формується при оптимальному мінеральному живленні рослин. При нестачі азоту і фосфору у живильному субстраті спостерігаються глибокі зміни ультраструктури хлоропластів, руйнуються або не утворюються тилакоїди, грани і міжгранальні ламели, з’являються пухирці й електроннощільні тіла. У хлоропластах клітин обкладкової паренхіми зменшуються розмір і кількість крохмальних зерен. При нестачі у живильному розчині азоту і фосфору структура хлоропластів мезофілу також зазнає змін.
Нестача, як калію і азоту та фосфору, порушує синтез хлорофілу. Зменшення вмісту пігментів у листках, структурні зміни у організації хлоропластів призводять до зниження інтенсивності фотосинтезу і у кінцевому результаті – до зменшення продуктивності рослин.
Зрозуміло, що крім перерахованих макроелементів, для нормального функціонування фотосинтетичного апарату необхідна низка інших макроелементів і мікроелементів.
Кисень. Концентрація його у атмосфері становить близько 21% і перевищує оптимальну для фотосинтезу величину. Підвищення і зменшення концентрації цього газу у повітрі позначається на фотосинтезі. У рослин з високим рівнем фотодихання (біб та ін.) зменшення концентрації кисню від 21 до 3 відсотків сприяло підвищенню фотосинтезу; у рослин з низьким фотодиханням (кукурудза) така зміна газового складу не мала суттєвого впливу на інтенсивність цього процесу.
Добовий і сезонний хід фотосинтезу. Серед факторів навколишнього середовища вирішальна роль за впливом на фотосинтез належить сумісній дії освітлення, температури і водного режиму. Ці фактори, особливо освітлення і температура, значно змінюють свої параметри протягом доби і викликають відповідні зміни у інтенсивності фотосинтезу (рис. 45).
В умовах помірного клімату максимальна інтенсивність спостерігається приблизно з 9-ої до 11-ої години дня. З підвищенням температури, зростанням водного дефіциту і зміною спектрального складу світла крива фотосинтезу спадає і може переміщуватися навіть нижче компенсаційної точки. У другій половині дня при поступовому зменшенні температури, що супроводжується, як правило, покращенням водозабезпечення листків рослин і частини червоного світла, інтенсивність фотосинтезу о 16-17-ій год знову зростає і досягає другого максимуму, який здебільшого менший від ранішнього. У північних широтах максимум інтенсивності фотосинтезу дещо зміщений у часі й крива одновершинна.

Рис. 45. Добовий хід фотосинтезу: 1 – у південних і середніх широтах;
2 – у північних широтах.
Нерівномірний хід фотосинтезу у онтогенезі рослин має відмінності, пов’язані з особливостями рослин, умовами кліматичної зони. За нормальних умов росту і розвитку максимум його припадає на період бутонізації – цвітіння рослин.
6.12. Генетика фотосинтезу. Міжвидова і внутрішньовидова
мінливість фотосинтезу
У вивченні проблеми фотосинтезу найбільше уваги на сьогодні надається вивченню генетичних відмін у активності фотосинтетичного апарату і пошуку шляхів її підвищення селекційно-генетичними способами. Дослідження фізіологічних характеристик культурних сортів різної продуктивності (переважно пшениці) та їхніх диких видів однозначно показали, що підвищення господарсько цінної продуктивності сільськогосподарських культур, досягнуте селекцією, відбулося не за рахунок активізації роботи фотосинтетичного апарату, а в основному внаслідок кращого розподілу асимілятів протягом онтогенезу і більш ефективного використання їх у процесі росту репродуктивних і запасаючих органів.
Швидкість фотосинтезу винятково стійка і слабко варіює у близькоспоріднених груп рослин. Існують генетично зумовлені особливості у організації енергосистем хлоропластів, які визначають можливу максимальну активність їх роботи.
Найбільш чіткі відміності інтенсивності фотосинтезу у С3 і С4-рослин.
Доведено, що фотосинтетичний апарат С3-видів у системі цілої рослини працює набагато менше своїх потенційних можливостей, тому що його активність обмежується попитом на асиміляти. Вимірювання світлових кривих фотосинтезу посівів різних культур при вирощуванні у оптимальних умовах показали, що швидкість поглинання СО2 при однакових характеристиках фотосинтетично активної радіації практично однакова для видів із С3-типом фотосинтезу. Для С4-рослин вона має свою закономірність (рис. 46).
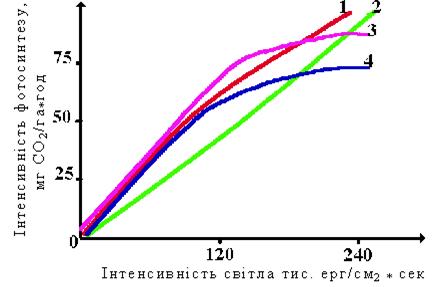
Рис. 46 - Світлові криві фотосинтезу різних культур:
1 – райграс; 2 – кукурудза; 3 – пшениця; 4 – картопля (за Гуляєвим Б. та ін. 1989).
Методи визначення інтенсивності фотосинтезу
Інтенсивність фотосинтезу характеризується кількістю СО2, що засвоюється одиницею листкової поверхні за одиницю часу. Звичайно цей показник виражають у мг СО2 на 1 дм2 за 1 год. Залежно від виду рослин і фази їх розвитку, інтенсивність фотосинтезу коливається у межах від 5 до 25 мг СО2/дм2×год.
Існує два визначення поняття фотосинтезу: дійсний і той, що спостерігається. Фотосинтез, що спостерігається, можна визначити за зменшенням концентрації СО2 або зростанням О2 в ізольованій камері, куди поміщено рослину. При визначенні дійсного фотосинтезу, крім визначення зміни газового складу ізольованої камери, визначають також витрати на дихання, тобто до знайденої експериментальної величини додають ту кількість СО2, яка могла б бути виділена листком у темряві. Але подібні розрахунки не наближають до оцінки “дійсного фотосинтезу” з таких причин: по-перше, при затіненні листка виключається не тільки “недійсний фотосинтез”, але й фотодихання; по-друге, так зване темнове дихання насправді залежить від світла.
Крім газометричних методів, існує багато інших, що дають можливість визначити інтенсивність засвоєння вуглецю рослинами. Серед них найбільш поширеними є радіометричне визначення фотосинтезу, визначення чистої продуктивності фотосинтезу, манометричний метод і деякі інші.
Фотосинтез як основа продуктивності
сільськогосподарських рослин
Фотосинтез є основним процесом утворення органічної речовини. У поєднанні з асиміляцією мінеральних елементів грунту він створює матеріальну базу для формування кількості і якості врожаю рослин. Про вирішальну роль фотосинтезу у формуванні врожаю свідчить те, що на частку органічної речовини припадає у середньому 95 відсотків маси сухої речовини тканин рослини. У молекулах органічних речовин зосереджений увесь запас енергії рослини.
Проте за сучасних технологій вирощування сільськогосподарських культур на створення відносно високих врожаїв використовують незначну кількість сонячної енергії: ярова пшениця – 3,26; картопля – 3,02; цукровий буряк – 2,12%.
Управління процесами фотосинтезу, їх регулювання є одним із найбільш ефективних шляхів управління продуктивністю рослин. Водночас необхідно враховувати, що загальна продуктивність рослинного організму залежить не тільки від інтенсивності фотосинтезу, але і від співвідношення між процесами асиміляції і дисиміляції, від ефективності використання органічних речовин, що виникли під час фотосинтезу, і від того, на які потреби рослина ці речовини переважно використовує.
В.І.Кефелі (1986) виділяє п’ять основних елементів продуктивності рослин, які регулюються генетико-гормональними факторами і перебувають у тісному взаємозв’язку (рис. 47).

Рис. 46. Елементи продукційного процесу.
Для автотрофних рослин ріст є первинним. Він забезпечує формування фотосинтетичних органів, починаючи від гетеротрофного розвитку, за рахунок запасних речовин насінини до активного здійснення фотосинтезу. Взаємовідношення фотосинтезу, дихання і росту при формуванні продуктивності перебувають під контролем генетичного фактора. Водночас ріст і розвиток пов’язані з живленням рослин макро- і мікроелементами. Під дією нітратів, що поглинаються рослинами, відбувається утворення цитокінінів, які транспортуються до стебел і листків й там беруть участь у клітинному поділі. Азотні сполуки беруть участь в утворенні ферментів, фосфорні – нуклеїнових кислот, макроенергетичних сполук та ін.
На хід процесів життєдіяльності впливає низка інших абіогенних факторів - низькі і високі температури, засолення, волога, радіація і т.п., на які рослина реагує відповідними реакціями. Це позначається на процесах росту і розвитку рослин, активності фотосинтетичного апарату, тривалості його функціонування.
Донорно-акцепторним зв’язкам належить основна роль у співвідношенні між процесами росту і розвитку, формуванні господарсько цінної продуктивності рослин.
Фотосинтезуючі органи рослин повинні забезпечувати органічними речовинами не тільки власні потреби, але і формування інших органів. У регулюванні розподілу асимілятів велика роль належить гормонам, зокрема абсцизовій кислоті. Вивчення фізіологічних характеристик культурних рослин та їхніх диких співродичів однозначно показало, що підвищення господарської продуктивності завдяки селекції відбувалося не за рахунок активізації роботи фотосинтетичного апарату, а головним чином унаслідок покращення стратегії розподілу асимілятів у рослині протягом онтогенезу і більш ефективного їх використання під час росту репродуктивних і запасаючих органів. Існує думка, що такий шлях у селекції значною мірою вичерпано і необхідно звернути увагу на добір форм рослин, що мають підвищеною активністю фотосинтетичного апарату. Водночас збільшення активності фотосинтетичного процесу є досить важким завданням у зв’язку з його складною генетичною зумовленістю та специфічністю ферментативних систем світлової і темнової стадій фотосинтезу.
6.15.Загальна біологічна продуктивність рослин
Кількість нагромадженої у процесі фотосинтезу речовини залежить від співвідношення між інтенсивністю фотосинтезу та інтенсивністю дихання. При досягненні рівноваги між інтенсивністю цих процесів спостерігається компенсаційна точка, яка найчастіше простежується при недостатньому освітленні рослин або при підвищеній температурі, що супроводжується дефіцитом води у рослині. Стан рослин, при якому виникає компенсаційна точка, є несприятливим для росту і накопичення врожаю. У рослини у такий період спостерігається стан бездіяльності – “простою”. При значному посиленні факторів, що викликають компенсаційну точку, крива фотосинтезу опускається нижче компенсаційної точки і створюється негативний баланс органічної речовини, тобто на дихання її витрачається більше, ніж синтезується. При усуненні цих факторів баланс стає знову позитивним.
Урожай рослин за певний період часу або за весь життєвий період є різницею між кількістю синтезованої і витраченої на дихання органічної речовини. Це є загальна продуктивність рослинного організму, або біологічний урожай, що визначається за формулою:
Уб = Іф - (Дт + Дф),
де: Уб – біологічний урожай; Іф – інтенсивність фотосинтезу; Дт – темнове дихання; Дф – фотодихання.
Практично чисту продуктивність фотосинтезу, що характеризує приріст органічної речовини за певний проміжок часу, віднесений до одиниці фотосинтезуючої поверхні, розраховують за формулою Кідда-Веста-Гріггса:
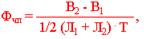
де: Фчп - чиста продуктивність фотосинтезу; В2 – В1 – приріст біомаси за проміжок часу Т; 1/2 (Л1 + Л2) - середня величина площі листків на початку і у кінці періоду визначення.
Ця величина непостійна і залежить від виду та сорту рослин, умов вирощування і коливається у межах 4–16 г/м2×добу.
Для людини більш цінною є та частина врожаю, що може бути використана як продукт харчування або сировина для промислової переробки. Вона називається господарсько цінною частиною врожаю.
Залежність між господарсько цінною частиною врожаю і біологічним урожаєм можна виявити з такого рівняння:
Уг = Уб – Мб,
де: Уг – урожай господарсько цінної частини врожаю; Уб – біологічний урожай;
Мб – урожай біологічної маси, що не використовується.
Якщо у це рівняння підставити значення Уб, то Уг дорівнюватиме:
Уг = Іф– (Дт + Дф + Мб).
З рівняння видно, що підвищити господарський урожай можна шляхом посилення фотосинтетичної діяльності рослини, зменшенням нераціональних витрат на дихання частини вегетативної маси, яка не використовується. У сучасних сортів пшениці й кукурудзи господарський урожай становить 0,5 біологічного.
Формування господарсько цінної частини врожаю відбувається головним чином у генеративний період. Тривале зберігання максимально високого рівня інтенсивності фотосинтезу у цей час є головним фактором продуктивності. Але дослідження свідчать про те, що часто на пізніх етапах спостерігається зниження фотосинтезу.
Останнім часом у питаннях підвищення продуктивності рослин покладаються надії на оптимізацію донорсько-акцепторних відношень між утворенням і використанням асимілятів у рослині.
Є підстави вважати, що конкуренція за асиміляти – досить поширене явище і властиве багатьом вищим рослинам. Досить активним споживачем асимілятів є коренева система. З появою нових споживачів, наприклад ранніх пагонів, може погіршитися забезпеченість коренів вуглеводами. Коренезабезпеченість (відношення маси коренів до наземної маси рослин) зменшується з часом росту і розвитку рослини. Важлива роль коренів у здійсненні регуляції фотосинтетичної функції добре ілюструється досить простими дослідами.
При видаленні частини кореневої системи гальмується фотосинтез. Гальмується інтенсивність фотосинтезу і при її охолодженні. При цьому надходження асимілятів до кореневої системи теж зменшується. Посилене надходження асимілятів до кореневої системи сприяє кращому забезпеченню фотосинтетичного апарату елементами мінерального живлення.
При формуванні генеративних органів виникає конкуренція за асиміляти, яка вирішується на користь генеративних органів, тому що стратегія рослин спрямована на забезпечення максимального коефіцієнта розмноження. Зменшити конкуренцію за асиміляти і цим подовжити фізіологічно активний стан рослин можна шляхом спрямованого керування розподілом асимілятів.
Завдяки селекції виведені більш продуктивні сорти, що мають велику господарсько цінну частину рослини. Це привело до різкого скорочення кількості асимілятів, які надходять до кореневої системи. Останнє може ще більше посилюватися в умовах високого агрофону, котрий, як відомо, сприяє насамперед відносному збільшенню наземної маси і зменшенню коренезабезпеченості рослин. Все це зменшує стійкість рослин до несприятливих факторів навколишнього середовища під час вегетації.
6.15.1. Світлокультура сільськогосподарських рослин
Інтенсивність світла і спектральний склад значною мірою визначають проходження окремих фаз росту і розвитку рослин. Практично регулювати інтенсивність і склад світла можна в умовах теплиць, оранжерей. У цих спорудах, крім того, піддаються регулюванню температура, мінеральне і повітряне живлення, водний режим. Вирощування рослин при штучному освітленні називається світлокультурою. Світлокультура дає можливість отримувати від 6 до 21 урожаїв овочевих культур за рік.
Спектральний склад світла впливає на якість врожаю. Якщо у складі світла переважають червоні промені, то у рослині синтезується більше білків. Ультрафіолетові промені у помірних дозах необхідні для нормального утворення органів рослин. У теплицях, парниках, накритих віконним склом, рослини не отримують необхідної кількості ультрафіолетових променів і мають тонкі й довгі стебла. Світло звичайних електричних ламп, що використовуються у теплицях, має мало фіолетових і багато жовто-червоних, особливо інфрачервоних, променів. До штучних джерел короткохвильового ультрафіолетового випромінювання належать електрична дуга і ртутні лампи у кварцовій оболонці – “Солюкс”, “Горное солнце” та ін. На сьогодні широко використовують люмінесцентні лампи, спектральний склад яких наближається до розсіяного денного сонячного світла. Ці лампи економічно дуже вигідні, оскільки перетворюють у світло 30–40% електричної енергії, а звичайні лампи розжарювання – лише 10%.
Сучасні споруди з регулюванням клімату мають різний ступінь складності. Найбільш прості з них – звичайні теплиці, у яких з відносною вірогідністю можна регулювати температуру, вологість повітря й умови освітлення рослин.
Більш складними є станції і лабораторії штучного клімату, які дозволяють більш чітко створювати певні параметри окремих елементів клімату.
Замкнутий комплекс або система споруд, що складається з декількох приміщень, у кожному з яких можна регулювати усі фактори клімату: температуру, вологу і склад повітря, інтенсивність і спектральний склад світла, добову тривалість освітлення, швидкість і напрямок руху повітря тощо, – має назву фітотрона. Перший фітотрон було споруджено у 1957 р. у Каліфорнійському технологічному інституті (США). Ефективність цього комплексу у вирішенні теоретичних і практичних проблем агрономії сприяла швидкому зростанню кількості фітотронів у різних країнах світу, у тому числі у нашій державі. В Україні фітотрони збудовані у Інституті генетики та селекції у Одесі та Інституті пшениці у Миронівці.
Це досить дорогі споруди, тому багато науково-дослідних і навчальних закладів користуються мікрофітотронами. Це пристрій, що складається з невеликої (від 1 до 5 м2) робочої камери вирощування рослин і машинної частини, якою створюється відповідний клімат. Мікрофітотрони називають нерідко вегетаційними шафами або камерами.
Важливими біологічними показниками умов освітлення, вирощування і фізіологічного стану рослин є анатомічна будова листків і стебел. Товщина листкової пластинки залежить від інтенсивності й спектрального складу світла. При застосуванні різних типів електричних ламп, навіть при однаковій активності опромінення, листки різних рослин мають неоднакову товщину. Найбільш товсті формуються під впливом люмінесцентних ламп. За анатомічними ознаками листків такі рослини можна віднести до групи світлолюбних. У них добре розвинені палісадна і губчаста тканини, число продихів на одиницю поверхні значно більше, ніж у листків, що виросли під лампами розжарювання. При цьому продихи є не тільки з нижнього боку листка, але зустрічаються і зверху.
Рослини краще поглинають світло, мають інтенсивніший газообмін, а отже і вищу продуктивність.
Листки рослин, що вирощуються взимку під лампами розжарювання, неоновими і ртутними лампами, за анатомічними ознаками належать до групи тіневитривалих рослин.
Спектральний склад штучного світла має значний вплив на ріст стебла і його діаметр. При довгохвильовому опроміненні під лампами розжарювання стебла подовжуються, а при короткохвильовому під люмінесцентними спостерігається укорочення міжвузлів.
Вплив штучного світла на інші фізіологічні показники вивчено недостатньо. Є низка дослідних даних про те, що синтез хлорофілу у більшості випадків швидше відбувається під впливом червоного, а каротиноїдів – синього світла.
6.15.2. Залежність фотосинтезу і газообміну фітоценозу від режиму ФАР
Знання природи фотосинтезу дозволяє орієнтуватися у виборі шляхів і методів впливу на цей процес з метою підвищення його ефективності. Серед цих шляхів важливе місце належить підвищенню коефіцієнта використання рослинами сонячної енергії. Середня величина цього коефіцієнта у рослин, що ростуть у природних умовах, досить низька і коливається у межах 0,5–1,5%. Підвищення його можливе за рахунок правильного розміщення рослин у посівах, збільшення площі листків до оптимальних розмірів і підвищення строку їх активної діяльності. Останнє досягається правильною системою удобрення, оптимізацією водного режиму, строками садіння та іншими агротехнічними заходами. Не менше значення має добір високопродуктивних сортів, найбільш придатних для конкретних грунтово-кліматичних умов.
Рослини можуть використовувати не весь спектр сонячного випромінювання, а лише ту частину, що має довжину хвилі 380–720 нм, тобто видиму частину спектра. Вона становить 26–55% загальної сонячної радіації. Чим вищий коефіцієнт використання фотосинтетично активної радіації (ФАР), тим вища врожайність сільськогосподарських культур.
О.А.Ничипорович вважає, що між коефіцієнтом корисної дії (к.к.д.) ФАР і врожайністю існує закономірність: при поглинанні 0,5–1,0% енергії ФАР урожайність є низькою; 1,1–2,0% – середня; 2,1-3,0% – добра; 3,1–4,0% - висока; 4,1–5,0% – дуже висока; 6,0–8,0% – теоретично можлива.
Якщо підняти к.к.д. культурних рослин від 1 до 3%, то у перерахунку на врожай зерна пшениці це становитиме 70–80 ц/га. Цим підвищенням використання енергії урожай сухої маси рослин на земній кулі зріс би на 20-25 млрд тонн, проти 7,5 млрд тонн що синтезуються нині всіма культурними фітоценозами.
У природних умовах фотосинтетично активна радіація міститься у прямому і розсіяному сонячному світлі. Прямим сонячним світлом називається та частина сонячного випромінювання, яка доходить до поверхні Землі у вигляді паралельних променів, що надходять безпосередньо від Сонця.
Розсіяною називається радіація, яка потрапляє на Землю після відбивання і розсіювання її молекулами газів повітря, твердими частками пилу, краплями води і крижинками льоду, що знаходяться у повітрі.
До сходу Сонця на рослини падає лише розсіяна радіація. Потім, при підніманні Сонця над горизонтом, частка прямої радіації відносно зростає, а розсіяної – швидко зменшується (рис. 48).

Рис. 47. Залежність прямої і розсіяної радіації від кута
падіння сонячних променів
Розсіяна радіація має велике значення у житті рослин. Близько 50–60% розсіяного світла становить фотосинтетично активна радіація.
Інтенсивність і спектральний склад сонячного світла залежать передусім від висоти Сонця над горизонтом. При низькому знаходженні у його випроміненні переважають інфрачервоні промені. Влітку, опівдні, у променях сонця, крім жовтих і червоних, міститься значна кількість синіх, фіолетових і ультрафіолетових променів.
Інтенсивність світла, як і його спектральний склад, залежать від нахилу Землі до Сонця, тобто від кута падіння сонячних променів. Цим кутом визначається і довжина шляху, яку проходить промінь через атмосферу Землі. Чим нижче сонце над горизонтом, тим менше сонячного випромінення доходить до поверхні Землі. Коли Сонце знаходиться на самому горизонті (0,5о), шлях сонячного променя у атмосфері буде у 35 разів довшим, ніж при розміщенні Сонця у зеніті.
Інтенсивність сонячного випромінювання, або кількість енергії, що припадає на 1 см2 за хвилину на межі атмосфери, становить близько 2кал/см2×хвилину. До земної поверхні енергії доходить значно менше, і визначається вона висотою Сонця над горизонтом і прозорістю атмосфери.
Сонячна енергія, що надходить до Землі, неоднаково взаємодіє з температурним режимом і вологою у різних географічних зонах. Внаслідок цього значно змінюється ефективність сонячної радіації як фактора продуктивності рослин.
6.15.3. Шляхи підвищення інтенсивності й продуктивності
фотосинтезу у посівах
Відносно взаємозв’язку площі асимілюючих органів з поглинанням енергії променів і продуктивністю рослин слід зазначити, що нині існує єдина думка: оптимальною для формування урожаю в основних сільськогосподарських культур є площа листків 30–40 тис.м2/га. Для характеристики асиміляційної поверхні використовується також термін “листковий індекс”, тобто відношення площі листків до площі посіву. Оптимальний індекс – 3–4, для окремих культур він становить 5–7.
Ефективність роботи фотосинтетичного апарату по створенню врожаю визначається не тільки листковим індексом, але й тривалістю життєдіяльності листків. Тому дуже важливо формувати посіви так, щоб якнайшвидше досягти оптимального листкового індексу і підтримувати його у такому стані досить тривалий час.
Фотосинтетичний потенціал – це один із вирішальних факторів, яким визначають величину врожаю, тому що він дає уявлення про те, яка фотосинтезуюча площа і протягом якого часу працювала на формування врожаю. Чим вищий фотосинтетичний потенціал (ФП), тим вища врожайність (якщо при цьому немає значного зменшення чистої продуктивності фотосинтезу).
Найвищі врожаї зерна ячменю отримують при ФП 2,6; озимої пшениці – 1,77; озимого жита – 1,5, більшості сортів картоплі - 1,8-2,1 млн.м2/добу.
Важливе значення для формування врожаю має структура посівів. Під цим поняттям розуміють штучно створювану архітектоніку агрофітоценозу, яка характеризується певними морфологічними ознаками і фізіологічними функціями. Оптимальною структурою вважається така, яка при високих к.к.д. фотосинтезу забезпечує максимальний біологічний і господарський урожай.
Листки різних ярусів рослин неоднаково поглинають сонячну енергію. Розподіл ФАР всередині посівів непропорційний площі листків окремих ярусів. Так, верхні листки, що становлять 23,7% всієї листкової поверхні рослини, поглинають 47% енергії. Листки середнього ярусу становлять 60,4% загальної площі, а поглинають лише 36,6% поглинутої посівами ФАР.
Підвищення фотосинтетичної продуктивності при оптимальному мінеральному живленні й водному режимі забезпечується найбільш раціональною архітектонікою, яка дає можливість рослинам досить ефективно засвоювати сонячну енергію. У сільськогосподарському виробництві це досягається різними строками і способами посіву з урахуванням напряму рядків і кількості рослин на одиницю площі. Кращі умови освітлення забезпечуються при розміщенні рядів посіву зі сходу на захід і з північного сходу на південний захід. Суттєве значення має селекція таких сортів, у яких листки рослин мають добре розвинену провідну систему, високоактивні ферментні системи асиміляції вуглекислого газу. Листки цих сортів повинні, крім того, відповідним чином розміщуватися на стеблах. В умовах високих географічних широт листки повинні розміщуватись під великим кутом відносно стебла. Косо спадаючі сонячні промені при такому розміщенні освітлюють усі яруси листків. У південних районах, навпаки, листки повинні розміщуватись під невеликим кутом до стебел з тим, щоб вертикально спадаючі промені досягли і найнижчих ярусів. Щоб не було взаємного затінення рослин і забезпечувалося добре освітлення листків усіх ярусів, суттєве значення має густота посівів.
Основними заходами по забезпеченню підвищення інтенсивності й продуктивності фотосинтезу у посівах є такі:
- селекція сортів з обмеженими ростовими процесами, сприятливою морфологічною структурою;
- підвищення родючості грунтів, створення умов для активної поглинальної діяльності кореневої системи (зменшення кислотності грунтів, їх щільності, оптимізація вологозабезпеченості та ін.);
- регулярне застосування органічних й сидеральних добрив з метою збільшення запасів органічної речовини у грунті і постійного надходження до приземного шару атмосфери СО2;
- своєчасне і достатнє забезпечення рослин необхідними елементами мінерального живлення, регуляторами росту;
- створення оптимальних умов освітлення посівів шляхом регулювання норм посіву, розміщенням у фітоценозах рослин різної морфологічної структури, оптимізації строків посіву та ін.;
- підтримання у активному стані асиміляційного апарату за допомогою агротехнічних і хімічних заходів боротьби із хворобами і шкідниками.
6.16. Кругообіг вуглекислого газу і кисню
Концентрація вуглекислого газу у атмосфері (0,032%) значно менша оптимального рівня, необхідного для повного насичення темнових реакцій фотосинтезу. Тому підвищення його концентрації до 0,3% і вище супроводжується зростанням інтенсивності й продуктивності фотосинтезу. При вмісті цього газу у повітрі 0,008% фотосинтез не відбувається.
У природних умовах рослини одержують СО2 не тільки з атмосфери, але й з грунту. За розрахунками Лундегорда, малогумусні піскові грунти за 1 год виділяють до 2 кг СО2 з кожного гектара, суглинкові грунти – близько 4 кг, а багаті на органічну речовину лісові грунти – від 10 до 25 кг. У середньому з 1га угноєного грунту виділяється щогодини близько 5 кг вуглекислого газу. Встановлено, що 1 га посіву вівса у процесі фотосинтезу споживає за 1 год 15 кг СО2. З них 5 кг забезпечує грунт і 10 кг – атмосфера, у шарі якої товщиною 100 м на площі 1 га міститься 550 кг вуглекислоти.
Зважаючи на те, що найбільш інтенсивно фотосинтез здійснюється протягом 8 год на добу, можна вважати, що 1 га посівів за 1 день споживає 120 кг СО2, з яких 40 кг надходить із грунту і 80 – з атмосфери. Отже, у денні години вміст вуглекислого газу у земному шарі повітря зменшується і відновлюється за решту годин доби. Баланс СО2 є позитивним на родючих грунтах і від’ємним – на бідних.
На рисунку 49 наведена схема кругообігу вуглекислого газу у природі.
Рослинність планети за дуже тривалий час свого існування поглинула з атмосфери величезну кількість СО2, надлишок якого створював “парниковий ефект”.
Щорічно зелені рослини споживають для фотосинтезу близько 3% запасів вуглекислого газу атмосфери і 0,3% його запасів у водах планети. Поглинання СО2 компенсується навіть із надлишком унаслідок зростаючих об’ємів спалювання пального, газу, вугілля, вирубування лісів, промислових викидів, вулканічної діяльності Землі та ін.
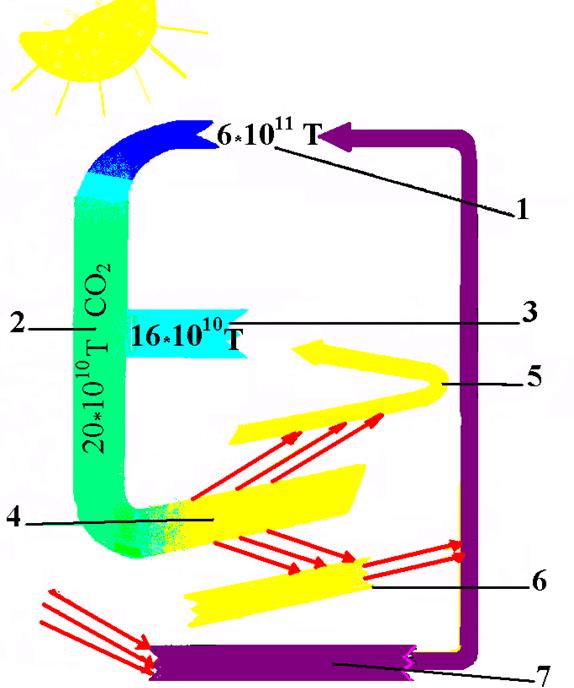
Рис. 49. Кругообіг вуглецю у природі (за Варасовою Н.М. і Шунтовою А.П.):
1 – вміст СО2 в атмосфері; 2 – засвоюється СО2 при фотосинтезі наземними рослинами;
3 – засвоюється рослинними організмами гідросфери; 4 – виділяється СО2 при перетворенні органічних речовин; 5- при диханні, 6 – перетворенні мертвих залишків;
7 – продуктів фотосинтезу минулих геологічних епох (вугілля, нафта, газ тощо).
Незважаючи на те, що кисень не бере безпосередню участь у фотосинтезі, він має значний вплив на цей процес. При збільшенні концентрації його у атмосфері й одночасному підвищенні освітленості у рослин виникає фотодихання.
Атмосфера має два джерела вільного кисню – дисоціацію молекул водяної пари під дією ультрафіолетових променів і фотосинтез. Перше джерело було, очевидно, єдиним постачальником кисню до появи на Землі хлорофілоносних рослин. Фотосинтез став основним джерелом масового накопичення кисню у атмосфері, вміст якого на сьогодні становить за об’ємом 21% і постійно підтримується на цьому рівні.
У нижніх шарах стратосфери під впливом коротких ультрафіолетових променів молекули кисню розпадаються на атоми, у зв’язку з чим стає можливим утворення молекули озону (О3). Більш довгі ультрафіолетові хвилі, що поглинаються озоном, руйнують його молекули. Залежно від поглинання енергії киснем і озоном, між ними встановлюється відповідне кількісне співвідношення. Область максимальної концентрації озону знаходиться на висоті 20–30 км. Тут створений своєрідний озоновий екран, яким поглинаються згубні для життя ультрафіолетові промені. Але значення озону полягає не тільки у цьому: він поглинає частину інфрачервоних променів, зменшуючи цим втрату тепла Землею.
Космічне значення рослин полягає насамперед у перетворенні сонячної енергії у потенційну хімічну, що нагромаджується у органічних сполуках кам’яного вугілля, нафти, газу, врожаю сільськогосподарських культур, маси лісів та ін. і вивільненні у атмосферу кисню, завдяки якому можливе життя аеробних організмів.
Питання для самоконтролю
Яке значення фотосинтезу для життя на Землі?
Які пігменти беруть участь у процесі фотосинтезу?
Від чого залежить зелений колір хлорофілу?
Чим пояснити флуоресценцію витяжки пігментів? Чому флуоресценція не спостерігається у живих листках?
Які промені поглинаються хлорофілами, каротиноїдами?
Охарактеризувати структурну формулу і хімічну природу хлорофілу.
Яка роль хлорофілу та каротиноїдів у фотосинтезі?
Суть світлової фази фотосинтезу. Що таке фотоліз води, фотосинтетичне фосфорилювання?
Суть темнової фази фотосинтезу.
Шляхи вуглецю при фотосинтезі. (С3–шлях, С4– лях, САМ–шлях).
Вплив інтенсивності світла на фотосинтез.
Особливості світлолюбних і тіневитривалих рослин.
Як змінюється спектральний склад сонячного світла протягом доби?
Добовий хід фотосинтезу.
Залежність інтенсивності фотосинтезу від температури.
Шляхи підвищення фотосинтезу в посівах.
Дата добавления: 2014-12-26; просмотров: 6257;