ГЛАВА 4. НЕРВНАЯ СИСТЕМА
Значение и функции нервной системы.Нервная система — система, объединяющая всю совокупность клеток нервной ткани в организме и вспомогательные структуры (оболочки мозга, сосуды и т. п.). Нервная система обеспечивает регуляцию всех жизненных процессов в организме и его взаимодействие с внешней средой, единство и целостность организма, а также формирует целенаправленное поведение.
Можно выделить два вида влияний нервной системы на органы и ткани: пусковое и моделирующее (корригирующее). Пусковое влияние вызывает деятельность органа, находящегося в покое. Прекращение импульсации, вызвавшей деятельность органа, ведет к возвращению его в исходное состояние. При поступлении импульсов к покоящейся скелетной мышце от мотонейронов спинного мозга или от мотонейронов ствола мозга по эфферентным нервным волокнам наступает ее сокращение. После прекращения импульсации в нервных волокнах, в частности в волокнах соматической нервной системы, сокращение мышцы также прекращается — мышца расслабляется.
Моделирующее (корригирующее) влияние.Этот вид влияния изменяет интенсивность деятельности органа. Оно распространяется как на органы, деятельность которых без нервных влияний невозможна, так и на органы, которые могут работать без пускового влияния нервной системы, то есть автономно. Примером моделирующего влияния на работающий орган может служить усиление или ослабление сокращения скелетной мышцы, а на органы, которые могут работать в автоматическом ритме — регуляция деятельности сердца, тонуса сосудов. Данное влияние может иметь разнонаправленный характер с помощью одного и того же нерва на разные органы. Так, моделирующее влияние блуждающего нерва на сердце выражается в угнетении его сокращений, но этот же нерв может оказывать пусковое влияние на пищеварительные железы, на находящуюся в покое гладкую мышцу желудка, тонкой кишки.
Моделирующее влияние осуществляется:
а) посредством изменения характера электрических процессов в возбудимых клетках органа возбуждения (деполяризация) или торможения (гиперполяризация);
б) за счет изменения кровоснабжения органа (сосудодвигательный эффект);
в) с помощью изменения интенсивности обмена веществ в органе (трофическое действие нервной системы).
Общий план строения нервной системы. Анатомически нервная система делится на центральную нервную систему (ЦНС) и периферическую нервную систему.
К первой относят головной и спинной мозг, вторая объединяет корешки, спинно-мозговые и черепные нервы, их ветви, сплетения и узлы, лежащие в различных отделах тела человека. По функциональному принципу нервную систему делят на анимальную (соматическую), иннервирующую собственно тело (сому), кроме внутренних органов, сосудов и желез; и вегетативную (автономную), регулирующую деятельность перечисленных органов.
Нервная система построена из двух типов клеток: нервных и глиальных, причем число последних примерно в 10 раз больше, чем нервных. Однако именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
Строение и функции нейрона.Нейрон — это структурно-функциональная единица нервной системы. Нейроны — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов. Особенностью нейрона является способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний — синапсов. Размеры нейронов колеблются от 6 до 120 мкм. В каждой нервной клетке выделяют четыре основных элемента (Атл., рис. 1, с. 118): тело или сому, дендриты, аксони пресинаптическое окончание аксона. Каждый из этих элементов выполняет определенную функцию.
От тела клетки берут начало дендриты и аксон. Дендриты ветвятся и на них расположены шипики, занимающие до 40% от поверхности сомы нейрона и дендритов. За счет шипиков воспринимающая поверхность нейрона значительно возрастает и может достигать, например, у клеток Пуркинье, 250 000 мкм2. Им принадлежит ведущая роль в восприятии информации. Если шипики не получают импульсации, то они исчезают. Сома и дендриты покрыты нервными окончаниями — синаптическими бутонами (бляшками) и отростками глиальных клеток. На одном нейроне число синаптических бляшек может достигать 10000. Аксон начинается от тела клетки аксонным холмиком, диаметр аксона составляет 1—6 мкм, на периферии длина аксона может достигать 1 м и более. Основной функцией аксона является проведение нервного импульса — потенциала действия. Аксон оканчивается утолщением — синаптической бляшкой, заполненной пузырьками, в которых хранятся химические вещества — медиаторы.
Тела нейронов составляют серое вещество мозга (ядра больших полушарий и мозжечка). Отростки нейронов составляют проводящие пути (белое вещество мозга). Мозг человека содержит около 25 млрд нервных клеток, взаимодействие между которыми осуществляется посредством синапсов, число которых в тысячи раз больше самих клеток (1015—1016), так как их аксоны многократно делятся. Нервные клетки имеются и вне ЦНС: периферический отдел вегетативной нервной системы, афферентные нейроны спинно-мозговых ганглиев и ганглиев черепных нервов. Периферических нервных клеток намного меньше, чем центральных — всего около 25 млн.
Классификация нейронов.По строению нейроны делят на три типа: униполярные, биполярные и мультиполярные (Атл., рис. 21, с. 17, рис. 2, с. 118).
Истинно униполярные нейроны находятся только в ядре тройничного нерва. Эти нейроны обеспечивают проприорецептивную чувствительность жевательных мышц. Главным образом эти нейроны характерны для нервной системы беспозвоночных. Другие униполярные нейроны называют псевдоуниполярными,и они имеют два отростка (один идет с периферии от рецептора, другой в структуры центральной нервной системы). Оба отростка сливаются вблизи тела клетки в единый отросток. Эти клетки располагаются в сенсорных узлах: спинальном, тройничном и т. д. Они обеспечивают восприятие болевой, температурной, тактильной, проприорецептивной, барорецептивной, вибрационной сигнализации.
Биполярные нейроны имеют один аксон и один дендрит. Нейроны этого типа встречаются в основном в периферических частях зрительной, слуховой и обонятельной систем. Биполярные нейроны дендритом связаны с рецептором, аксоном — с нейроном следующего уровня организации соответствующей сенсорной системы.
Мультиполярные нейроны имеют несколько дендритов и один аксон. В настоящее время насчитывают до 60 различных вариантов строения мультиполярных нейронов, однако все они представляют разновидности веретенообразных, звездчатых, корзинчатых и пирамидных клеток.
По чувствительности к действию раздражителей нейроны делят на моно-, би-, полисенсорные. Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей сенсорности. Например, значительная часть нейронов первичной зоны зрительной области коры реагирует только на световое раздражение сетчатки глаза.
Бисенсорные нейроны.Чаще располагаются во вторичных зонах коры кого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности. Например, нейроны вторичной зрительной коры больших полушарий реагируют на зрительные и слуховые раздражители.
Полисенсорные нейроны.Это нейроны ассоциативных зон мозга; они способны реагировать на раздражение слуховой, кожной и других рецептивных систем.
Функционально нейроны делятся на три типа: афферентные, интернейроны (вставочные), эфферентные.
Афферентные нейроны — нейроны, воспринимающие информацию с помощью рецепторов о внешней и внутренней среде организма и передающие ее в центральную нервную систему. Вступая в пределы ЦНС, окончания их отростков устанавливают синаптические контакты со вставочными, а иногда и непосредственно с эфферентными нейронами.
Вставочные нейроны, или интернейроны, обрабатывают информацию, получаемую от афферентных нейронов, и передают ее на другие вставочные или на эфферентные нейроны. Область влияния вставочных нейронов определяется их собственным строением, и, как правило, они имеют аксоны, разветвления которых заканчиваются на нейронах своего же центра, обеспечивая, прежде всего, их интеграцию. Одни вставочные нейроны получают активацию от нейронов других центров и затем распространяют эту информацию на нейроны своего центра. Это дает усиление влияния сигнала за счет его повторения в параллельных путях и удлиняет время сохранения информации в центре. В итоге центр, куда пришел сигнал, повышает надежность воздействия на исполнительную структуру.
Другие вставочные нейроны получают активацию от коллатералей эфферентных нейронов своего же центра и затем передают информацию назад в свой же центр, образуя обратные связи. Так организуются реверберирующие сети, позволяющие длительно сохранять информацию в нервном центре.
Эфферентные нейроны нервной системы — это нейроны, передающие информацию от нервного центра к исполнительным органам или другим центрам нервной системы. Так, например, эфферентные нейроны двигательной зоны коры больших полушарий — пирамидные клетки, посылают импульсы к мотонейронам передних рогов спинного мозга, то есть они являются эфферентными для этого отдела коры большого мозга. В свою очередь мотонейроны спинного мозга являются эфферентными для его передних рогов и посылают сигналы к мышцам. Основной особенностью эфферентных нейронов является наличие длинного аксона, обладающего большой скоростью проведения возбуждения.
По активности различают фоноактивные и «молчащие нейроны», возбуждающиеся только в ответ на раздражение. Фоноактивные нейроны имеют большое значение в поддержании тонуса нервных центров, уровня возбуждения коры и других структур мозга. Число фоноактивных нейронов увеличивается в состоянии бодрствования.
Важную роль в деятельности нервной системы играют глиальные клетки. Глиальные клетки (нейроглия — «нервный клей») более многочисленны, чем нейроны, составляют около 50% от объема ЦНС. Они способны к делению в течение всей жизни. По размеру глиальные клетки в 3—4 раза меньше нервных, их число достигает 14 x 1010, с возрастом их количество увеличивается (число нейронов уменьшается). Глиальные клетки выполняют несколько функций: опорную, защитную, изолирующую, обменную (снабжение нейронов питательными веществами). Микроглиальные клетки способны к фагоцитозу, ритмическому изменению своего объема (период «сокращения» — 1,5 мин, «расслабления» — 4 мин). Циклы изменения объема повторяются через каждые 2—20 ч. Предполагают, что пульсация способствует продвижению аксоплазмы в нейронах и влияет на ток межклеточной жидкости. Мембранный потенциал клеток нейроглии составляет 70—90 мВ, однако потенциал действия не генерируют, генерируют только локальные токи, распространяющиеся от одной клетки к другой. Следовательно, глиальные клетки способны к передаче возбуждения, распространение которого от одной клетки к другой идет с декрементом. Распространению возбуждения между глиальными клетками способствуют специальные щелевые контакты их мембран. Эти контакты обладают пониженным сопротивлением и создают условия для электротонического распространения тока от одной глиальной клетки к другой. Процессы возбуждения в нейронах и электрические явления в глиальных клетках, по-видимому, взаимодействуют (Смирнов, 2000).
Различают несколько видов нейроглии, каждая из которых образована клетками определенного типа: астроцитами, олигодендроцитами, микроглиоцитами (Атл., рис. 22, с. 17).
Астроциты представляют собой многоотростчатые клетки с ядрами овальной формы и небольшим количеством хроматина. Размеры астроцитов 7—25 мкм. Астроциты располагаются главным образом в сером веществе мозга. Ядра астроцитов содержат ДНК, протоплазма имеет пластинчатый комплекс, центрисому, митохондрии. Считают, что астроциты служат опорой нейронов, обеспечивают репаративные процессы нервных стволов, изолируют нервное волокно, участвуют в метаболизме нейронов. Отростки астроцитов образуют «ножки», окутывающие капилляры, практически полностью покрывая их. В итоге между нейронами и капиллярами располагаются только астроциты. Видимо, они обеспечивают транспорт веществ из крови в нейрон и обратно (Покровский и др., 2001). Астроциты образуют мостики между капиллярами и эпендимой, выстилающей полости желудочков мозга. Считают, что таким образом обеспечивается обмен между кровью и цереброспинальной жидкостью желудочков мозга, то есть астроциты выполняют транспортную функцию.
Олигодендроциты — клетки, имеющие малое количество отростков. Они меньше по размеру, чем астроциты. В коре больших полушарий количество олигодендроцитов возрастает от верхних слоев к нижним. В подкорковых структурах, в стволе мозга олигодендроцитов больше, чем в коре. Олигодендроциты участвуют в миелинизации аксонов (поэтому их больше в белом веществе мозга), в метаболизме нейронов, а также в трофике нейронов.
Микроглия представлена самыми мелкими многоотростчатыми клетками. Источником микроглии служит мезодерма. Микроглиальные клетки способны к фагоцитозу.
Основные и специфические функции нервных клеток. Основные функции нейрона идентичны общим функциям любых клеток организма.
1. Синтез тканевых и клеточных структур, а также необходимых для жизнедеятельности соединений (анаболизм). При этом энергия не только расходуется, но и накапливается, поскольку клетка усваивает органические соединения, богатые энергией (белки, жиры и углеводы, поступающие в организм с пищей). В клетку питательные вещества поступают в виде моносахаров, аминокислот, жирных кислот и моноглицеридов. Процесс синтеза обеспечивает восстановление структур, подвергающихся распаду.
2. Выработка энергии в результате катаболизма — совокупности процессов распада клеточных и тканевых структур и сложных соединений, содержащих энергию. Энергия необходима для обеспечения жизнедеятельности клетки.
3. Трансмембранный перенос веществ, обеспечивающий поступление в клетку необходимых веществ и выделение из клетки метаболитов и веществ, используемых другими клетками организма.
Специфическими функциями нейрона являются:
1. Восприятие изменений внешней и внутренней среды организма. Данная функция осуществляется с помощью периферических нервных образований — сенсорных рецепторов и посредством шипикового аппарата дендритов и тела нейрона.
2. Передача сигнала другим нервным клеткам и клеткам рабочих органов: скелетной мускулатуры, гладким мышцам внутренних органов, сосудам, секреторным клеткам. Эта передача реализуется с помощью синапсов.
3. Переработка поступающей к нейрону информации, пришедшей к нему в виде нервных импульсов.
4. Хранение информации с помощью механизмов памяти. Любой сигнал внешней или внутренней среды организма преобразуется в процесс возбуждения, который является наиболее характерным проявлением активности любой нервной клетки.
5. Нервные импульсы обеспечивают связь между всеми клетками организма и регуляцию их функций.
6. С помощью химических веществ нервные клетки оказывают трофическое веяние на эффекторные клетки организма (питание).
Общая физиология нейрона. Основные понятия.Нервная клетка как мышечные и секреторные клетки, относится к возбудимым клеткам и обладает рядом свойств: раздражимостью, возбудимостью, проводимостью. Раздражимость — общее свойство всех живых клеток.
Раздражимость — это способность клеток отвечать на внешние или внутренние воздействия изменением своего состояния. Частным проявлением раздражимости является возбудимость.
Возбудимость — это способность тканей при действии раздражителей переходить в активное состояние. Нейрон способен генерировать потенциал действия (ПД) на раздражение. ПД проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток.
Раздражитель— это любое изменение внешней или внутренней среды организма. По видам энергии раздражители делят на физические (электрические, механические, температурные, световые), химические, биологические и социальные (для человека). В зависимости от степени чувствительности клеток к тому или иному раздражителю — на адекватные и неадекватные. Адекватный раздражитель — это такой раздражитель, к которому клетка наиболее чувствительна и к его восприятию приспособлена в процессе эволюции, например, свет для фоторецепторов глаза, звук — для слуховых рецепторов, пахучие вещества — для обонятельных и т. д. Адекватным раздражителем для нейронов и мышечных клеток, многоклеточных организмов являются некоторые химические вещества и электрический ток. Все прочие раздражители называют неадекватными. Например, механическое воздействие на фоторецепторы или нервные волокна.
Нервная клетка может находиться в двух состояниях: пассивном и активном. Пассивное,или физиологический покой, — это состояние относительного покоя, отсутствие раздражителей, когда обменные процессы направлены на поддержание жизнедеятельности клетки. Активное состояние, как указывается в специальной физиологической литературе, проявляется процессом возбуждения и торможения.
Возбуждение — активное состояние клетки, ткани, которое возникает при действии раздражителей и проявляется в специфической деятельности (например, сокращении мышцы, генерации электрического сигнала, выброса секрета).
Торможение — активное состояние, возникающее при действии раздражителей или идущее вслед за возбуждением, проявляется в прекращении деятельности клетки, ткани (например, уменьшение метаболизма, снижение возбудимости по отношению к раздражителям).
Проводимость — способность ткани и клетки проводить возбуждение. Проводимость нервной ткани связана с распространением в ней процессов возбуждения. Основным условием для проведения возбуждения является целостность мембраны.
Биоэлектрические явления в нервных клетках. Для исследования электрических явлений в нервных (и других) клетках в настоящее время используют микроэлектроды (стеклянные пипетки с очень тонким кончиком 0,5 мкм), заполненные электролитом, который играет роль проводника тока, а стекло — изолятора. Микроэлектрод вводят внутрь клетки, и он регистрирует внутриклеточный потенциал (относительно наружного «индифферентного» электрода). С помощью внутриклеточных электродов удалось зарегистрировать электрические ответы возбудимых тканей.
1. Потенциал покоя. У нейронов, как и других возбудимых тканях, поверхностная мембрана в покое электрически поляризована, то есть имеет разный электрический потенциал между наружной и внутренней сторонами. Его величина в нервных клетках составляет 60—80 мВ относительного нулевого потенциала внешней среды. Эту величину называют потенциалом покоя (ПП). Во всех точках аксоплазмы этот потенциал практически одинаков. При регистрации ПП луч осциллографа во время прокола мембраны клетки микроэлектродом отклоняется и показывает отрицательный заряд внутри клетки.
Согласно мембранно-ионной теории (Берштейн, Ходжкин, Хаксли, Катц), непосредственной причиной формирования ПП является неодинаковая концентрация анионов и катионов внутри и вне клетки (Атл., рис. 5, с. 120). В нейронах концентрация ионов К+ внутри клетки в 20—40 раз больше, чем вне клетки, концентрация ионов Nа+ вне клетки в 10—12 раз больше, чем в клетке. Ионов Сl- вне клетки в 10—20 раз больше, чем внутри клетки. Различные ионы распределены неравномерно по обе стороны клеточной мембраны, во-первых, вследствие неодинаковой проницаемости клеточной мембраны для различных ионов, во-вторых, в результате работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки концентрационному и электрическому градиентам.
Мембрана легко проницаема для жирорастворимых, веществ, молекулы которых проникают через слой липидов, являющихся матриксом мембраны. В мембране нервного волокна имеются каналы, проницаемые для воды, ионов Nа+, К+, Cl–, Ca2+. При этом К+ из клетки выходит в значительно большем количестве, чем входит Na+ в клетку, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости для Na+.
Органические анионы из-за своих больших размеров не могут выходить из клетки, поэтому внутри клетки в состоянии покоя отрицательных ионов оказывается больше, чем положительных. Поэтому клетка внутри имеет отрицательный заряд. Калий является главным ионом, обеспечивающим формирование ПП. В состоянии покоя клетки устанавливается динамическое равновесие между числом выходящих из клетки и входящих в клетку ионов К+.
Потенциал покоя — это алгебраическая сумма не только всех зарядов ионов вне и внутри клетки, но также алгебраическая сумма отрицательных внешних и внутренних поверхностных зарядов самой мембраны, и составляет величину порядка 60—80 мВ.
Кроме собственно ионных механизмов формирования ПП, связанных с различной проницаемостью клеточной мембраны и диффузией ионов, имеется активный механизм поддержания градиентов концентрации различных ионов внутри и вне клетки. Их называют натрий-калиевой помпой.
2. Потенциал действия. Потенциал действия (ПД), или потенциал возбуждения, нервных клеток (волокон) возникает в ответ на достаточное по силе раздражение. Потенциал действия — кратковременный электрический процесс, выражающийся в быстром колебании мембранного потенциала вследствие перемещения ионов в клетку и из клетки и способный распространяться без затухания (без декремента). Он обеспечивает передачу сигналов между нервными клетками, между нервными центрами и рабочими органами.
В процессе развития ПД возникают фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое движение иона Nа+ в клетку, а иона К+ из клетки. Величина мембранного потенциала сначала уменьшается, по абсолютной величине до нуля, а затем снова восстанавливается до исходного уровня. Уменьшение мембранного потенциала называют деполяризацией. Когда деполяризация достигает примерно 50% пороговой величины, возрастает проницаемость ее мембраны для ионов Nа+, но скорость сравнительно невелика. Движущей силой, обеспечивающей вход ионов Nа+ в клетку являются концентрационный и электрический градиенты. При возбуждении нейрона повышается проницаемость его мембраны и для ионов Са+, но его ток в клетку значительно меньше, чем ионов Nа+. Условием, обеспечивающим вход иона Nа+ вклетку, и последующий выход иона К+ из клетки, является увеличение клеточной проницаемости клеточной мембраны, которая определяется состоянием воротного механизма ионных Nа+-и К+-каналов. Воротный механизм Nа+-каналов расположен на внешней стороне клеточной мембраны (Nа+движется внутрь клетки), воротный механизм К+-каналов — на внутренней (К+ движется из клетки наружу).
Активация Nа+- и К+-каналов (открытие ворот) обеспечивается уменьшением мембранного потенциала. Когда деполяризация клетки достигает критической величины, которая обычно составляет 50 мВ, проницаемость мембраны для ионов Nа+ резко возрастает — открывается большое число ворот Nа+- каналов, и ионы Nа+ лавиной устремляются в клетку. В результате интенсивного тока ионов Nа+ внутрь клетки далее процесс деполяризации проходит очень быстро, в итоге ПП исчезает, становится равным нулю. Фаза деполяризации на этом заканчивается.
При увеличении силы раздражителя и достижении порога раздражения, то есть критического уровня деполяризации, возникает потенциал действия.
В потенциале действия различают пик (спайк) и следовые потенциалы. Пик ПД представляет собойкратковременную инверсию, происходит перезарядка мембраны, за счет продолжающегося входа Nа+ в клетку (ворота Nа+-каналов еще открыты). Поэтому число положительных ионов в клетке превосходит число отрицательных, заряд внутри клетки становится положительным, снаружи — отрицательным (атл., рис. 5, с. 120). В фазе инверсии электрический градиент препятствует входу Nа+ внутрь клетки (положительные заряды отталкиваются друг от друга), проводимость Nа+ снижается, однако некоторый период (доли миллисекунды) ионы Nа+ продолжают входить в клетку, о чем свидетельствует продолжающееся нарастание ПД. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом ионов Nа+ в клетку.
Примерно через 0,5—1 мс после начала деполяризации рост ПД прекращается, так как закрываются ворота Nа+-каналов и открываются ворота К+-каналов, то есть увеличивается проницаемость для ионов К+. Поскольку ионы К+ находятся преимущественно внутри клетки, то согласно концентрационному градиенту они быстро выходят из клетки, вследствие чего в клетке уменьшается число положительно заряженные ионов. Заряд клетки начинает возвращаться к исходному уровню. В фазе инверсии выходу ионов К+ из клетки способствует также электрический градиент. Ионы К+ выталкиваются положительным зарядом из клетки и притягиваются отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки — до конца фазы инверсии, когда начинается следующая фаза ПД — фаза реполяризации. Калий выходит из клетки по управляемым каналам и каналам утечки.
В фазе реполяризации проницаемость клеточной мембраны для ионов К+ все еще высока, и они продолжают быстро выходить из клетки согласно концентрационному градиенту. Клетка снова внутри имеет отрицательный заряд, а снаружи — положительный (Атл., рис. 5, с. 120), поэтому электрический градиент препятствует выходу К+ из клетки, хотя он продолжает выходить. Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее действия электрического градиента. Таким образом, вся нисходящая часть пика ПД обусловлена выходом иона К+ из клетки вследствие закрытия ворот К+-каналов.
3. Следовые потенциалы выражаются в гиперполяризации или частичной деполяризации клетки после возвращения мембранного потенциала к исходной величине. Следовые потенциалы имеют значительно меньшую амплитуду (их величина составляет 5—10% от высоты пика). Различают следовую деполяризацию и следовую гиперполяризацию.
Следовой деполяризацией называют следовой отрицательный потенциал — это длительное сохранение отрицательности наружной поверхности возбужденного участка к невозбужденному, в этот период наружная поверхность возбудимого образования имеет больший отрицательный заряд, чем в состоянии покоя. Следовая деполяризация обусловлена кратковременным повышением проницаемости клеточной мембраны для Nа+ и входом его в клетку согласно концентрационному и электрическому градиентам, мембрана еще сохраняет отрицательный заряд.
Следовой гиперполяризацией называют следовой положительный потенциал, что соответствует периоду увеличения мембранного потенциала покоя, то есть гиперполяризации мембраны. В этот период поверхность клетки более положительно заряжена, чем в состоянии покоя. Это связано с повышенной проницаемостью мембраны для ионов К+. Ворота К+-каналов еще не полностью закрыты, поэтому К+ продолжает выходить из клетки согласно концентрационному градиенту, что ведет к гиперполяризации клеточной мембраны, после чего происходит полное восстановление потенциала покоя. Таким образом, волна возбуждения состоит из потенциала покоя (порогового потенциала), потенциала действия (или пика потенциала действия), следовой деполяризации и следовой гиперполяризации. В нервных волокнах млекопитающих, имеющих наибольший диаметр, длительность пика ПД составляет 0,4—0,5 мс, следовой отрицательности — 12—20 мс и следовой положительности — 40—60 мс.
Изменение возбудимости клетки при возбуждении. Возбудимость клетки во время ее возбуждения не остается постоянной, а изменяется. Выделяют следующие фазы изменения возбудимости.
1. Кратковременное повышение возбудимости отмечается в начале развития ПД, когда уже возникла частичная деполяризация клеточной мембраны. Возбудимость повышена потому, что клетка частично деполяризована, мембранный потенциал приближается к критическому уровню, поскольку открывается часть Nа+-каналов.
2. Абсолютная рефрактерная фаза, то есть полная невозбудимость клетки (возбудимость равна нулю), соответствует пику ПД и продолжается 1—2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефрактерная фаза. Клетка в этот период при любой силе раздражителя не отвечают. Невозбудимость клетки в фазу деполяризации и инверсии (восходящая часть пика ПД) объясняется тем, что Nа+-каналы открыты и ионы Nа+ быстро поступают по всем каналам. Под влиянием деполяризации продолжают открываться те ворота Nа+-каналов, которые еще не успели открыться. Поэтому дополнительное раздражение клетки относительно движения ионов Nа+ в клетку ничего изменить не может. Именно поэтому ПД либо совсем не возникает на раздражение, если оно мало, либо возникает максимальным, если оно достаточной силы (пороговой или сверхпороговой). В период нисходящей части инверсии и реполяризации клетка также невозбудима, потому что закрываются Nа+-каналы, в результате чего клеточная мембрана непроницаема для ионов Nа+, даже при сильном раздражении. Кроме того, в этот период открываются К+-каналы, К+ быстро выходит из клетки, обеспечивая нисходящую часть фазы инверсии и реполяризацию. Абсолютная рефрактерная фаза в процессе реполяризации продолжается до момента, когда мембранный потенциал будет примерно на критическом уровне деполяризации. В это время около половины Nа+-каналов возвращается в исходное состояние, поэтому возможна их новая активация. Абсолютный рефрактерный период ограничивает максимальную частоту генерации ПД. Если абсолютный рефрактерный период завершается через 2 мс после начала ПД, то клетка может возбуждаться с частотой максимум 500 имп/с. Существуют клетки с еще более коротким рефрактерным периодом, в которых возбуждение может повторяться с частотой 1000 имп/с. Так, например, клетки ретикулярной формации центральной нервной системы.
3. Относительная рефрактерная фаза — это период восстановления возбудимости, когда сильное раздражение может вызвать новое возбуждение, она продолжается 4—8 мс и соответствует следовой отрицательности клеточной мембраны, что является следствием все еще повышенной проницаемости для ионов К+ и избыточного выхода ионов К+ из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить сильное раздражение, так как часть каналов в конце реполяризации находится еще в состоянии инактивации, а выход ионов К+ из клетки препятствует ее деполяризации.
4. Фаза экзальтации (супернормальность) — то есть повышенной возбудимости, появляющейся от следовой деполяризации. В этой фазе очередной ПД можно вызвать более слабым (подпороговым) раздражителем, поскольку мембранный потенциал несколько ниже обычного, в связи с повышенной проницаемостью клеточной мембраны для ионов Nа+. Длительность этой фазы 10—15 мс.
5. Субнормальность, то есть пониженная возбудимость, возникающая от следовой гиперполяризации, длительность в пределах 20—50 мс. Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность.
Лабильность, или функциональная подвижность(Н. Е. Введенский), — это скорость протекания электрических и биохимических процессов, обеспечивающих функциональную деятельность ткани. Исходя из определения, лабильность ткани зависит от длительности ПД. Это означает, что лабильность, какПД, определяется скоростью перемещения ионов в клетку и из клетки, которая, в свою очередь, зависит от скорости изменения проницаемости клеточной мембраны. Особое значение при этом имеет длительность рефрактерной фазы: чем больше рефрактерная фаза, тем ниже лабильность ткани.
Мерой лабильности является максимальное число потенциалов действия, которое способна генерировать ткань в 1 с в точном соответствии с ритмом раздражения. Различные ткани обладают разной степенью лабильности. Так, например, наибольшей лабильностью отличаются аксоны, они воспроизводят до 500—1000 импульсов в секунду; менее лабильны синапсы (центральные и периферические места контакта), способны передавать не более 100—150 возбуждений в секунду. Наиболее лабильными являются волокна слухового нерва, в которых частота генерации потенциалов действия достигает 1000 Гц.Лабильность клеток понижается при длительном бездействии, при утомлении.
Следует отметить, что при постепенном увеличении частоты ритмического раздражения лабильность ткани повышается, то есть ткань отвечает более высокой частотой возбуждения по сравнению с исходной частотой. Однако в максимальном темпе ткань работать не может и поэтому наряду с максимальным ритмом раздражения для характеристики лабильности в физиологии предложен оптимальныйтемп. Показателем оптимального ритма является частота раздражения, которая в течение длительного времени воспроизводится без трансформации. Оптимальный ритм является собственно «рабочим» ритмом возбудимого образования. Условия для длительного воспроизведения ритма создаются при частоте, когда каждый следующий импульс падает в период повышенной возбудимости, то есть в фазу экзальтации.
Пессимальный ритм — чрезмерная частота (выше максимальной), при которой ткань отвечает не на каждый ритм, то есть часть раздражений попадает в фазу рефрактерности.
Оценка возбудимости нервной ткани и клетки. Возбудимость клетки меняется не только в процессе ее возбуждения, атакже в результате длительной активности. Показателями состояния возбудимости ткани являются: а) пороговая сила и б) пороговое время.
Пороговая сила — это наименьшая сила раздражителя, способная вызвать возбуждение (ПД). Чем меньше его величина, тем больше возбудимость. Силу раздражителя меньше порога называют подпороговой, больше порога — надпороговой. Сила раздражителя, вызывающая самую большую реакцию, называется максимальной силой. Сила раздражителя больше порога, но меньше максимальной называется субмаксимальной, а сила, превышающая максимальную, — супермаксимальной. Все раздражители, дающие максимальные ответы, называют оптимальными. Силы больше по величине, чем оптимальные, но вызывающие меньший ответ, чем при оптимальном раздражении, называют пессимальными. При использовании в качестве раздражителя электрического тока предложенное определение пороговой силы совпадает с понятием «реобаза». Реобаза — это наименьшая сила тока, способная вызвать возбуждение. Если пороговая сила раздражителя мала, возбудимость ткани высокая. Чем ниже пороговая сила, тем выше возбудимость ткани. Большая пороговая сила свидетельствует о низкой возбудимости ткани.
Пороговое время — это минимальное время, в течение которого на ткань должен действовать раздражитель пороговой силы, чтобы вызвать ее возбуждение. Пороговое время называют также полезным временем, так как дальнейшее увеличение раздражения уже становится бесполезным. По его величине можно судить о функциональном состоянии нервной ткани. Для оценки возбудимости ткани чаще используют не пороговое время, а хронаксию. Хронаксия — это наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение. В течение этого времени происходит уменьшение мембранного потенциала до величины, соответствующей критическому уровню деполяризации. Соотношение между временем действия раздражителя и пороговой силой выражают кривой (Гоорвег — Вейса — Лапика), она носит название кривая «силы — времени» (рис. 8).
| напряжение тока, В Б А |
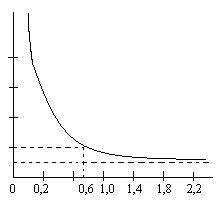
| время, мс |
| Б |
| А |
| б |
| а |
Рис. 8. Кривая «силы — времени»: А — порог (реобаза);
Дата добавления: 2017-11-04; просмотров: 819;