ЗНАЧЕНИЕ ПОЛИПЛОИДИИ В СЕЛЕКЦИИ ДРЕВЕСНЫХ РАСТЕНИЙ
Широкое распространение полиплоидии у растений и важность этого биологического явления в эволюции обусловлены повышенной мощностью и лучшими адаптационными возможностями полиплоидных организмов. Эта особенность полиплоидии в сочетании с гибридизацией открывает большие перспективы в селекции растений. Значение полиплоидии в селекции растений трудно переоценить.
1. Полиплоидия позволяет преодолеть нескрещиваемость при гибридизации, что значительно расширяет возможности отдаленной межвидовой и даже межродовой гибридизации.
2. На полиплоидном уровне стерильные отдаленные гибриды становятся фертильными, это дает возможность вести дальнейшую селекцию гибридных форм у растений, неспособных к вегетативному размножению.
3. Благодаря полиплоидии становится возможным закрепление гетерозиса, когда два аллеля, обусловливающие гетерозис включаются в геном как два отдельных генома, в результате чего особи, имеющие генотип АВ АВ, будут обладать преимуществами гетерозиготного состояния и в то же время не будут производить в потомстве гомозиготные особи.
4. Полиплоидия и отдаленная гибридизация позволяют выявить видовые и родовые генетические потенциалы изменчивости, что очень важно для селекционной оценки исходного материала и выбора методов селекции.
Большой интерес для селекции растений представляют агамные поли-плоидные комплексы, состоящие из половых и апомиктических форм. Благодаря сочетанию полиплоидии, гибридизации и апомиксиса агамные полиплоидные комплексы представляют собой преуспевающие растительные группы, для которых характерен огромный полиморфизм и способность к расширению их ареалов даже за счет экологических ниш с неблагоприятными условиями].
Агамные полиплоидные комплексы образуются обычно у многолетних растений, способных к вегетативному размножению. Такие комплексы, например, описаны в роде Rubus у европейских и американских ежевик, у мяты, у вероник.
У лесных древесных растений агамные полиплоидные комплексы не описаны. Однако у видов, для которых характерны сложные полиплоидные ряды с нечетным числом геномов, можно предположить наличие апомиксиса и образование агамных комплексов, например у Morus nigra L., Acer rubrum L., Salix glauca L. и др. Выявление и изучение агамных полиплоидных комплексов у лесных древесных растений позволит использоватьихкак ценный исходный материал для дальнейшей синтетической селекции.
В селекции на гетерозис спонтанная и экспериментальная полиплоидия в сочетании с гибридизацией широко используются у самых разных сельскохозяйственных культур: пшеница, рожь, сахарная свекла, табак, хлопчатник, картофель, арбузы и др. В селекции цветочных и декоративных растений также используется гибридизация и экспериментальная полиплоидия. Большие успехи в селекции плодовых, цитрусовых и ягодных растений достигнуты благодаря использованию отдаленной гибридизации, спонтанной и экспериментальной полиплоидии.
Спонтанная и экспериментальная полиплоидия в сочетании с гибридизацией широко применяется в селекции шелковицы.
Лесные древесные виды в пределах большинства родов характеризуются слабой хромосомной дифференциацией, поэтому проблема нескрещиваемости при межвидовой гибридизации для большинства видов не стоит так остро, как у многих травянистых растений. С этой точки зрения использование экспериментальной полиплоидии в селекции лесных древесных растений не может иметь большого значения. Лишь в отдельных частных случаях, когда генетические барьеры нескрещиваемости между видами значительно выражены (например, у некоторых видов сосны), искусственную полиплоидию, возможно, будет целесообразно использовать для преодоления нескрещиваемости.
При отдаленных межвидовых скрещиваниях диплоидных видов у лесных древесных получены интересные гибриды, сочетающие некоторые ценные признаки родительских форм, например у елей и у сосен. Возникшую при этом стерильность гибридов теоретически возможно преодолеть с помощью искусственной полиплоидизации, но это требует экспериментального доказательства и разработки соответствующих методик для лесных древесных растений.
Экспериментальная полиплоидия у древесных использовалась в селекции декоративных форм у сосны, дуба, тополя.
В селекции на соматический и адаптивный гетерозис у лесных растений этот способ нельзя считать перспективным, так как искусственная полиплоидия, как правило, генетически не сбалансирована и не обладает гомеостатичностыо.
Наиболее перспективным в селекции лесных древесных растений следует считать использование спонтанной аллополиплоидии и искусственной гибридизации, особенно для вегетативно размножаемых пород,
На примере родов Populus и Salix нами показано, что скрещиваемость и возможность получить гетерозис в этих родах различны: у тополей на диплоидном уровне скрещиваемость зависит от степени филогенетического родства: чем ближе скрещиваемые виды, тем лучше их скрещиваемость и больше шансов получить гетерозис; у ив при скрещивании даже самых филогенетически далеких разнохромосомных видов, принадлежащих к разным секциям и подродам, можно получить высокую скрещиваемость и гетерозис.
Сопоставляя данные кариологических исследований и изучения гетерозиса у гибридов-сибсов ивы белой и ивы ломкой (табл. 10) можно отметить такую тенденцию: с увеличением уровня плоидности степень выраженности гетерозиса увеличивается.
Коэффициент гетерозиса определялся по Нансону и Сакре по формуле Н% = 2АВ 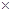 100 : (А + В) - 100, где Н% - коэффициент гетерозиса, АВ - показатели гибридов, А и В - показатели родительских форм.
100 : (А + В) - 100, где Н% - коэффициент гетерозиса, АВ - показатели гибридов, А и В - показатели родительских форм.
Закономерности скрещиваемости и выраженности гетерозиса, выявленные при скрещивании видов ив с разным числом хромосом, могут быть использованы и при межвидовой гибридизации в родах Betula; Alnus и других древесных растений, в которых эволюция кариотипов шла по пути естественной полиплоидизации.
Дата добавления: 2017-08-01; просмотров: 2466;