ОБОСНОВАНИЕ МЕТОДОВ СЕЛЕКЦИИ
Определив направление селекции, необходимо решить целесообразность применения тех или иных методов селекции (отбор, селекция географических экотипов, гибридизация, полиплоидия, мутагенез и др.) или сочетание этих методов.
В популяционной селекции для создания устойчивых лесных биоценозов целесообразно использование методов массового отбора лучших географических экотипов, внутривидовой межпопуляционной гибридизации различных экотипов и изолятов; возможно также использование межвидовой интрогрессивной гибридизации.
Задача селекции – получить генетически сбалансированные популяции с высокой генетической изменчивостью, обладающие популяционным или групповым гетерозисом.
В селекции, целью которой является создание высокопродуктивных интенсивных культур с коротким оборотом рубки, целесообразно преимущественно использование методов отдаленной гибридизации, полиплоидии с последующим индивидуальным отбором.
Задача этого направления селекции – получить синтетические смеси или отдельные клоны, обладающие индивидуальным соматическим гетерозисом.
В селекции для защитного лесоразведения целесообразно использование индивидуального и массового отбора, гибридизации и полиплоидии.
В задачу этого направления селекции входит получение синтетических популяций, обладающих групповым или индивидуальным гетерозисом главным образом адаптивным.
Использование вышеописанных теоретических предпосылок разработки селекционных программ улучшения лесных древесных можно рассмотреть на примере селекции конкретных лесных пород.
Для хвойных пород характерна медленная адаптивная эволюция, в результате которой преобладает географический тип видообразования и формирование больших панмиктических популяций. Для реликтовых видов характерны небольшие гомозиготные по многим признакам изолированные популяции. Полиплоидия у хвойных – явление очень редкое. Способность к инбридингу у хвойных выражена довольно четко, но индивидуальные различия в пределах видов значительны. Репродуктивная хромосомная изоляция в разных родах выражена по-разному. Наиболее строгая изоляция характерна для рода Pinus. Этим, видимо, объясняется отсутствие доказательных примеров интрогрессивной гибридизации у сосен.
При искусственной межвидовой гибридизации у сосен и елей получен групповой и индивидуальный гетерозис, но эта форма гетерозиса генетически несбалансирована. Такие гибриды хуже приспосабливаются к условиям среды, чем природные популяции. Поэтому в популяционной селекции искусственная межвидовая гибридизация у елей и сосен не может быть главным стратегическим направлением, большие результаты в генетическом улучшении этих пород можно получить при внутривидовой межпопуляционной гибридизации.
В роде Picea широко распространена межвидовая интрогрессивная гибридизация, так как хромосомная изоляция видов в этом роде слабо выражена, генетические барьеры нескрещиваемости видов почти отсутствуют.
При разработке способов вегетативного размножения использование естественных гибридов в качестве исходного материала – один из перспективных методов в селекции елей для плантационных насаждений.
В роде Larix механизмы репродуктивной изоляции видов также выражены очень слабо, а в некоторых случаях отсутствуют полностью,
Таблица
Примерная схема целесообразных направлений и методов селекции основных древесных видов в б. СССР
| Метод селекции | Необходимые мероприятия | Вид | Популяция | |
| Популяционная селекция для создания лесных биоценозов | ||||
| 1. Массовый отбор | Отбор лучших происхождений Pinus sylvestris, | Генетически | ||
| в географических опытах | Picea abies, Picea Obovata, Quercus Robur, Larix sibiri- сa, Larix europea | Сбалансированные панмиктические Популяции в центре ареалов | ||
| 2. Внутривидовые | Создание семенных планта ций из лучших географиче ских происхождений с вы сокой ОКС с целью полу чения семян для выращива ния синтетических популя ций, обладающих высоким гомеостазом и популяцион- ным гетерозисом | Тоже | Тоже | |
| Межпопуляцион- ные, географиче ски отдаленные скрещивания (свободное опы ление) | ||||
| 3. Внутривидовые межпопуляцион- ные скрещивания двух изолятов а) свободное опыление б) искусственное Опыление | а) Подбор изолятов с высокой ОКС. Создание прививочных семенных плантаций из двух изолятов и их свободное опыление | Picea fennica | Изолированные малые популя- ции | |
| Picea orientalis | ||||
| Pinus sylvestris | ||||
| Larix kurilensis, | ||||
| Larix lubarskii | ||||
| б) Сбор пыльцы в одном изоляте и опыление ею деревьев другого изолята выращивание синтетиче ских популяций, обладающих попупяционным гетерозисом | Larix olgensis Betula maxirno- wicziana, Betula pendula, Betula Pubescens | |||
| Межвидовая гибридизация в комбинациях, дающих групповой гетерозис (искусственные скрещивания и интрогрессивная гибридизация | Скрещивание индивидов с высокой СКС и выращивание синтетических гибридных популяций с групповым гетерозисом | Populu” alba X X P. tremula, La rix leptolepis X X L. europea. Quercus robur X X Q. Petrea Picea abies X X P. Obovata | Популяция на границах ареалов | |
| На границах больших и малых популяций | ||||
Селекция для плантационных интенсивных культур
| Внутривидовая, межпопуляционная гибридизация свободное опыление) |
| Отбор родительских деревьев с высокой СКС, их клонирование черенками или прививками, создание семенных садов для получения семян с гетерозисным эффектом |
| Pinus sylvestris, Picea abies, P. obovata, Larix europea, L. sibirica |
| Тоже |
| Таблица (окончание) | ||||
| Межвидовая гибридизация (искусственная) | Отбор деревьев с высокой СКС, скрещивание, индиви дуальный отбор гибридов, их проверка и вегетативное размножение | Populus, Salix, Ксеа abies X Р. obo vata,Larix sibirica, L. europea, L. Lepto Lepis | Малые изолированные популяции | |
| Интрогрессивная гибридизация (свободное опыление) | Отбор естественных гибридов и их вегетативное размножение | Populus suaveolens X P. Laurifolia, P. amurensis X X P. Maximowiczii | Популяции на границах больших перекрещивающихся ареалов | |
| Отдаленная гибридизация разно- Хромосомных аллоплоидных видов (искусственное скрещивание) | Подбор пар и видов с разным Salix alba 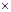 S. tra- S. tra-
| Популяции в экстремальных условиях на границах ареалов или в изолятах | ||
| чистом хромосом, скрещива- gilis, Betula pendula | ||||
| ние, индивидуальный отбор гибридов на гетерозис и декоративность, их вегетативное размножение | 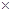 P. Pubescens, Ainus glutinosa P. Pubescens, Ainus glutinosa 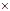 A. hirsuta A. hirsuta
| |||
| Селекция защитного лесоразведения Селекция для защитного Лесоразведения | ||||
| Межвидовая гибридизация искусственная и интрогрессивная | Скрещивание деревьев с высокой ОКС и СКС. Выращивание синтетических гибридных популяций с даптивным | Quercus robus Q. Petrea, Populus nigra P. Deltojdes
| Популяции на границах больших ареалов | |
| адаптивным | ||||
| групповым гетерозисом | ||||
| Отдаленная гибридизация разнохромосомных аллоплоидных видов | Межвидовое и межсекционные скрещивания видов с разным числом хромосом. Отбор гибридных комбинаций с групповым гетерозисом и индивидуальный отбор гетероэисных форм | Betula pendula B. pubescens, Betula pendula B. papyrifera
Ainus glutinosa A. hirsuta, A. incana. Salix alba S. fragffis
| Популяции в экстремальных условиях на границах ареалов и изолятов |
результатом чего является широкое распространение интрогрессивной гибридизации у лиственниц, В этом роде одним из ведущих стратегических направлений в селекции для плантационных насаждений наряду с использованием интрогрессивной гибридизации может стать искусственная межвидовая гибридизация. Многочисленные исследования межвидовых гибридов, особенно европейской и японской лиственниц, показали возможность получения группового гетерозиса.
У лиственных пород наряду с географическим типом видообразования может встречаться быстрое квантовое видообразование, особенно при переопылении полуколониальных и колониальных изолятов. В эволюции многих родов лиственных пород большую роль сыграла полиплоидия. Поэтому пути эволюции лиственных более многообразны, чем у хвойных.
В роде Quercus доказаны широкие возможности межвидовой гибридизации и получения индивидуального гетерозиса, но его использование проблематично из-за отсутствия надежных способов вегетативного размножения дуба. Следует далее разрабатывать возможность использования интрогрессивных межвидовых гибридов дуба. Для популяционной селекции наиболее целесообразны методы внутривидовой межпопуляционной гибридизации различных географических экотипов и отбор лучших природных популяций.
В селекции быстрорастущих пород в родах Populus и Eucalyptus получены ценные гибриды при межвидовой гибридизации.
Способность к вегетативному размножению позволяет использовать индивидуальный гетерозис гибридов тополей и эвкалиптов при их плантационном выращивании.
В родах, эволюция которых шла с участием полиплоидии (Salix, Morus, Betula и др.), наиболее перспективна селекция на гетерозис, при которой отдаленная гибридизация в сочетании с полиплоидией позволяет получить и групповой, и индивидуальный гетерозис, при этом у гибридов наблюдается огромный формовой полиморфизм, охватывающий весь спектр мутационной изменчивости, характерной для этих родов.
В табл. приведена примерная схема определения целесообразных направлений и методов селекции лесных древесных видов в зависимости от их генетико-эволюционных особенностей и целей селекции.
ЗАКЛЮЧЕНИЕ
Накопленный в селекции древесных опыт показал, что генетическое улучшение лесных древесных невозможно без знания генетико-эволю-ционных характеристик исходного материала.
Общебиологические закономерности эволюционного процесса растений характеризуются следующей схемой.
В процессе эволюции естественных популяций и видов мутационный процесс, поток генов между популяциями и рекомбинации аллелей создают изменчивость генотипов, а естественный отбор и дрейф генов сортируют эту изменчивость.
Естественный отбор проявляется в различных формах. В стабильных популяциях, хорошо приспособленных к определенным условиям, отбор элиминирует "крайние" отклонения. Эта форма отбора, называемая стабилизирующей, наиболее распространена в природе и характерна для популяций многих древесных растений. Часто стабилизирующий или уравновешивающий отбор благоприятствует гетерозиготам, в результате такого отбора создается сбалансированный полиморфизм, основанный на превосходстве гетерозигот. Примером могут служить некоторые популяции сосны обыкновенной.
Дизруптивный отбор благоприятствует сохранению "крайних" типов и элиминации промежуточных. Эта форма отбора усиливает полиморфизм, примером могут служить популяции древесных и стланниковых форм высокогорных сосен и ив на границе субальпийской и альпийской зон.
Направляющий или прогрессивный отбор вызывает направленное в одну сторону изменение генетического состава популяции при освоении новой среды.
Соотношение главных движущих сил эволюции их взаимодействие в разных ситуациях будет различным, в зависимости от этого пути и скорость эволюции будут различаться. В больших панмиктических популяциях эффективность естественного отбора очень высока, а роль генетического дрейфа незначительна; эти популяции высокогомеостатичны, эволюция в них протекает медленно и адаптивно. В малых изолированных популяциях, где панмиксия нарушена, естественный отбор мало эффективен, в популяции господствуют случайные колебания мутаций, эволюция в них протекает быстро, но не адаптивно. В таких популяциях в результате инбридинга и дрейфа генов создается большой полиморфизм. При нарушении изоляции поток генов между такими полиморфными популяциями поставляет материал для возникновения гетерозиса и быстрой адаптивной эволюции, поскольку становится возможным сочетание потока генов, рекомбинаций, отбора и генетического дрейфа в их тесном взаимодействии. На границах видовых ареалов в условиях интрогрессии в результате потока генов и рекомбинации возникает большая изменчивость. У лесных древесных растений явление интрогрессивной гибридизации известно широко и характерно для многих лесообразующих видов (ели, лиственницы, березы, дуба и др.). Гибридные популяции часто занимают огромные площади, особенно в нарушенных антропогенным влиянием местообитаниях, где создаются экологические ниши для распространения гибридных межвидовых рекомбинатов.
Большую роль в эволюции новых форм играет полиплоидия, являющаяся наиболее эффективным средством достижения целесообразных форм дупликаций генов. Совместное действие гибридизации и полиплоидии создает совершенно новые аллоплоидные виды, для которых характерно определенное сочетание признаков, часто отличных от родительских форм.
Многообразие путей эволюции формирует различные группы растений (популяции, виды, роды), различающиеся по характеру генетико-эволю-ционных процессов и по закономерностям формообразовательного процесса.
В общей схеме эволюционного процесса лесные древесные растения имеют свои особенности.
Для всех древесных с их длительным жизненным циклом характерна медленная адаптивная эволюция. Географический тип видообразования обычно преобладает, однако при восстановлении потока генов между полуколониальными и колониальными горными и островными изолята-ми возникают случаи быстрого квантового видообразования, у лиственных пород этот тип видообразования встречается чаще, чем у хвойных.
Все лесные древесные растения – анемофильные и энтомофильные перекрестнопылители, обладающие в известной мере системами самонесовместимости. Однако способность к инбридингу у разных видов различна – от самофертильных растений (Picea omorica) до частично самостерильных (виды родов Pinus, Picea, Abies). У частично самостерильных видов хорошо выражены индивидуальные различия по реакции на самоопыление. Инбредная депрессия тоже различна у разных видов, индивидуумов, степень ее выраженности зависит и от условий выращивания.
Хромосомная дифференциация видов в пределах родов древесных слабо выражена. Однако механизмы репродуктивной изоляции видов в разных родах различны. Наиболее строгая хромосомная изоляция характерна для многих видов рода Pinus, в родах Picea и Larix у большинства видов репродуктивная хромосомная изоляция почти отсутствует; у лиственных пород генетические барьеры межвидовой несовместимости выражены слабо. Не скрещиваются лишь виды, филогенетически далекие.
В связи с этими особенностями хромосомной дифференциации у древесных на границах ареалов, при их перекрывают и совпадении сроков цветения часто наблюдается интрогрессивная гибридизация.
Несмотря на определенное однообразие морфологии хромосом в пределах родов, все же имеются некоторые различия между популяциями и даже в пределах популяции по общей длине хромосом, распределению перетяжек и др. По характеру этой изменчивости можно в известной мере судить о направлении эволюционного процесса в изучаемой популяции.
Эволюция кариотипа у растений вообще и у древесных в частности идет двумя путями:
а) хромосомные перестройки,
б) полиплоидизация.
Первый путь характерен для видов с крупными хромосомами и небольшими хромосомными числами, сюда относятся большинство хвойных пород, второй – для видов с мелкими хромосомами. В эволюции многих родов и видов лиственных пород полиплоидия сыграла важную роль, поэтому пути их развития более многообразны, чем у хвойных.
Анализ генетико-эволюционных особенностей исходного материала позволяет выбрать целесообразные пути и методы генетического улучшения лесных древесных растений и проводить селекцию в известной мере целенаправленно.
Для определения оптимальной селекционной стратегии необходимо последовательное выявление видовых, географических, популяционных и индивидуальных особенностей и. их связи с селектируемыми признаками в исходном материале.
На популяционных моделях определяется тип отбора, аддитивная ценность, характер наследования селектируемых признаков, их возрастная и экологическая изменчивость, корреляционные связи признаков, компоненты продуктивности.
В популяциях, сформировавшихся под действием стабилизирующего отбора, с явно выраженным взаимодействием генотип–среда у видов, скорость роста которых меняется в онтогенезе, например Pinus sylvestris, Picea abies, индивидуальный отбор генотипов на повышение продуктивности малоперспективен. Больший селекционный эффект у этих видов можно получить при выявлении географических и межпопуляционных различий.
Для интенсивных плантационных насаждений с сокращенным ротационным циклом эффективность географической и популяционной селекции может быть повышена за счет селекции клонов, селекции на СКС межвидовой гибридизации с последующим кленовым отбором. Для ели обыкновенной в ряде зарубежных стран эти методы уже апробированы. Для правильной оценки результатов селекции и выбора направления необходима дальнейшая разработка методов генетического анализа количественных признаков.
Классический генетический анализ, базирующийся на гибридологическом анализе, в большинстве случаев не применим при изучении количественных признаков растений. Генетический анализ количественных признаков подразумевает в первую очередь анализ наследственной обусловленности наблюдаемого разнообразия в популяциях растений.
Из всего арсенала методов количественной генетики растений при изучении древесных пород наибольшие возможности применения имеют методы разграничения компонент общей изменчивости, оценки комбинационных способностей и выявления степени взаимосвязи различных признаков.
Средиметодов разграничениякомпонент изменчивости и определения наследуемости признаков из традиционных способов при работе с древесными могут быть использованы методы дисперсионного и регрессионного анализов. Особый интерес представляют методы, не требующие смены поколений. Однако большинство из них не применимы из-за низкой достоверности получаемых результатов. Наиболее теоретически обоснованным и приспособленным к нуждам селекции является метод фоновых признаков, разработанный В.А. Драгавцевым и П.С. Дьяковым, но и он требует доработки в отношении применимости конкретно к древесным растениям. Перспективным может оказаться также метод сопоставления лимитов изменчивости в клонах и популяции, предложенный В.М. Роне.
Знание величин эффектов общей и специфической комбинационной способностей важно как при разработке общей стратегии селекции отдельных культур, так и при оценке конкретных генотипов на разных стадиях отбора. В зависимости от необходимой степени точности оценки искомых параметров может быть применен тот или иной метод анализа (поликросс, топкросс или диаллельный анализ). Диаллельный анализ дает возможность получить наиболее точную информацию, но вместе с тем он значительно более трудоемок. При работе с древесными применима схема анализа данных диаллельного скрещивания по Гриффингу, но с учетом того, что оцениваемые параметры могут быть в некоторой степени искажены из-за гетерозиготности родительских форм и возможного наличия эпистаза. Необходимо учитывать также, что на результаты диаллельного анализа сильное влияние имеют эффекты взаимодействия генотипа и среды.
Оценка степени взаимосвязи признаков подразумевает в первую очередь определение коэффициентов корреляции на фенотипическом, генотипическом и паратипическом уровне. Однако с увеличением числа признаков резко возрастает число коэффициентов, они становятся труднообозримыми и нелегко поддаются интерпретации. В таких случаях большую помощь могут оказать такие методы многомерного статистического анализа, как факторный анализ и анализ главных компонент. Эти методы позволяют выделить основные факторы, контролирующие изученные признаки и показать степень взаимосязи факторов и признаков.
Один из важных путей генетического улучшения древесных — гибридизация. В разных странах накоплен большой экспериментальный материал по гибридизации основных лесообразующих видов.
Межвидовая гибридизация усиливает формообразовательный процесс. В результате комбинаций генов при объединении разных геномов возникают совершенно новые формы растений, особенно при сочетании гибридизации с полиплоидией. В комбинациях скрещиваний родительских форм с высокой СКС можно получить гетерозис, использование которого открывает большие перспективы в повышении продуктивности лесных древесных. Однако гетерозис получается в большинстве случаев не в гибридной семье в целом, а лишь у отдельных индивидуумов. Поэтому для видов, способы вегетативного размножения которых еще не разработаны (виды родов Quercus, Pinus и др.) использование межвидовой гибридизации ограничено. Для видов легко размножающихся вегетативно (тополи, ивы, эвкалипты) межвидовая гибридизация с последующим клоновым отбором очень перспективна для плантационных культур интенсивного типа.
В отдельных случаях установлена возможность получения группового гетерозиса в гибридных семьях, например при скрещивании Larix europea L. leptolepis, Dopulus tremula P.tremuloides, Populus tremula P. alba. Межвидовая гибридизация в этих комбинациях может быть использована не только в клоновой, но и в популяционной селекции для создания высокопродуктивных синтетических популяций. Однако следует учитывать, что при межвидовой гибридизации часто получается генетически несбалансированный гетерозис типа пышности по Добжанскому, поэтому гибридные синтетики могут быть менее гомеостатичными, чем естественные популяции.
Внутривидовая межпопуляционная гибридизация и гибридизация географических экотипов в последнее время показала большие перспективы в популяционной селекции сосен, елей, лиственниц. В этом случае возможно получение популяционного гетерозиса и создание устойчивых синтетических популяций.
Возможности гибридизации расширяются в связи с использованием полиплоидии. Полиплоидия позволяет преодолеть нескрещиваемость при отдаленной гибридизации, что дает возможность получить гибриды филогенетических далеких видов, вплоть до межподродовых. На поли-плоидном уровне отдаленные гибриды могут быть фертильными, что позволяет вести дальнейшую селекцию у растений, неспособных к вегетативному размножению.
Благодаря полиплоидии становится возможным закрепление гетерозиса в последующих поколениях. И наконец, отдаленная гибридизация полиплоидов позволяет выявить видовые и родовые потенциалы генетической изменчивости, что очень важно для селекционной оценки исходного материала и выбора методов селекции.
Так как обнаруженная при скрещиваниях разнохромосомных видов изменчивость подчиняется закону гомологических рядов Н.И. Вавилова, становится возможным прогнозирование новых форм гибридов. У лесных древесных видов в родах Salix, Ainus, Betula широко распространена спонтанная полиплоидия. Использование отдаленной гибридизации в сочетании с полиплоидией в этих родах имеет большие перспективы.
С гибридизацией и полиплоидией тесно связана селекция на гетерозис. Это одно из ведущих направлений в селекции.
Гетерозис – очень сложное биологическое явление, которое нельзя ограничивать только понятием "гетерозиготность" и связывать его лишь с первым поколением, так как практически гетерозис можно получить не только при скрещивании чистых линий, но и при отдаленной гибридизации, а на тюлиплоидном уровне гетерозис часто сохраняется во втором и последующих поколениях.
Для выяснения механизма гетерозиса предлагается расчленить понятие "гетерозис" на более простые составляющие.
По биологической сущности нами выделены три категории гетерозиса:
1 – популяционный гетерозис, формирующийся в естественных популя- циях в процессе эволюции под действием уравновешивающего отбора;
2 – групповой гетерозис — получается при искусственной или интро-грессивной гибридизации родительских форм с высокой СКС;
3 – индивидуальный гетерозис также возникает при искусственной и интрогрессивной гибридизации, когда в гибридных комбинациях лишь отдаленные индивидуумы обладают гетерозисом.
По всем трем категориям выделяются типы гетерозиса по характеру его проявления – репродуктивный, соматический, адаптивный; и по типу взаимодействия генов – сверхдоминирование, комплементация, аддитивные эффекты.
На клоновых биологических моделях, состоящих из гибридов с различным типом гетерозиса и их родительских форм проведены многоплановые комплексные исследования на организменном, тканевом, клеточном и молекулярном уровнях.
Исследованы составляющие биомассы, анатомическая структура листьев, ультраструктура хлоропластов, термоустойчивость клеток листа, цитологическая характеристика активности меристем, особенности нуклеинового обмена, степень гетерогенности изоферментов.
Генетико-статистический анализ изменчивости изученных признаков показал большое генетическое разнообразие клонов, включенных в биологическую модель гетерозиса. Выявлены взаимосвязи признаков, что, с одной стороны, позволило упростить анализ за счет удаления неинформативных признаков, а с другой – дало возможность обосновать подходы для прогнозирования гетерозиса. Показано изменение характера взаимосвязи в системе признаков продуктивности в зависимости от отсутствия или присутствия гетерозиса и его типа.
Эти исследования позволили выявить особенности проявления различных типов индивидуального гетерозиса. Некоторые из этих особенностей раскрывают и общебиологические закономерности в сложных механизмах гетерозиса.
Дальнейшая интенсификация селекционного процесса у лесных древесных в плане исследований эффектов популяционного, группового и индивидуального гетерозиса, а также полиплоидии связана с необходимостью расширения исследований природы гетерозиса и полиплоидии на всех уровнях организации растений – молекулярном, клеточном, тканевом, органном, организменном, популяционном. При этом следует иметь в виду, что результаты исследований на низших иерархических уровнях лишь частично могут быть экстраполированы на высшие уровни. Не случайно, например, одним из основных предметов генетики растений является изучение признаков продуктивности у отдельных растений, а продуктивность популяций и ценозов изучается биологами с использованием всего арсенала знаний различных областей современной биологии – генетики, физиологии, биохимии, цитологии, биофизики и других на разных уровнях организации жизнедеятельности. Даже сам генетический анализ растительных объектов от молекулярного уровня до популяционного предусматривает использование принципов и подходов как молекулярной генетики, так и генетики популяций.
Результаты современных генетических и биохимических исследований показывают, что в основе явлений гетерозиса, полиплоидии и депрессии растений лежат определенные перестройки в молекулярной структуре генома и плазмона. Об этом свидетельствуют, в частности, различия между родительскими формами гетерозисных гибридов растений по содержанию повторяющихся нуклеотидных последовательностей ДНК, а также увеличение относительного содержания повторов ДНК у полиплоидных видов растений. Эти вариации в структуре геномов растений могут быть определены с использованием методов дифференциальной окраски хромосом, кинетики реассоциации ДНК, гибридизации индивидуальных фракций ДНК и отдельных типов РНК с ДНК. Как видно из этого, методы молекулярной биологии и генетики дополняют классические способы выявления хромосомных перестроек, обусловленных дупликациями, делениями, транслокациями и инверсиями.
В функциональном отношении перестройки структуры хромосом в областях локализации повторов ДНК при инбредной депрессии, гетерозисе и полиплоидии выявляются по степени деконденсации хромосом, ускорению репликации и транскрипции ДНК и повышению активности трансляционного аппарата. Эти функции хромосом осуществляются при активном участии регуляторных генов и участков ДНК, а также гормонально-ингибиторного комплекса, являющихся основными компонентами систем интеграции метаболизма, дифференцировки и морфогенеза растений.
Одним из важных механизмов активизации продукционного процесса у растений является интенсификация процессов репликации ДНК на разных этапах онтогенеза растений. У высокогетерозисных гибридов это проявляется главным образом в повышении активности митозов в корнях и побегах на начальных этапах онтогенеза и в зародышах сразу после процесса оплодотворения. Это преимущество на начальных этапах формирования вегетативных и репродуктивных органов выражается на поздних этапах онтогенеза в виде заметного увеличения биомассы соответствующих органов. Наряду с этим у высокопродуктивных форм растений интенсификация ростовых процессов в отдельных дифференцированных тканях обеспечивается также благодаря явлению клеточной полиплоидии.
Высокая продуктивность растений сопряжена с активацией синтеза РНК и белка. Повышение транскрипционной активности заметно в области ядрышкового организатора, т.е. в районе расположения генов рРНК. Повышение активности биосинтеза белка при гетерозисе происходит в основном за счет комплементации функциональной активности всех компонентов и сопровождается ускорением биосинтеза полипептидной цепи за единицу времени. Следовательно, эффект комплементации при гетерозисе может иметь место на уровне трансляции.
Принято считать, что наследование хозяйственно-полезных признаков растений связано с полигенным комплексом генов. В этой связи представляет интерес случай, когда состав популяций новообразованных иРНК родительских форм гетерозисных гибридов растений существенно отличается. В основном эти различия связаны с неодинаковой частотой встречаемости в популяции иРНК фракций, составляющих ее значительную часть. Последнее выявляется при гибридизации новообразованных РНК с ДНК.
Важное место среди подходов по изучению природы гетерозиса занимает изучение явления биохимического полиморфизма. Исследованиями многих авторов показана адаптивная роль полиморфизма, и поэтому анализ популяций, а также исходных форм высокопродуктивных гибридов с использованием электрофоретических методов позволяет достаточно обоснованно прогнозировать эффекты как генетического, так и онтогенетического гомеостаза. В этом плане необходимы исследования по подбору комплекса маркерных белков и ферментов, индекс разнообразия которых в конечном итоге будет определять в известной мере эффективность реализации селекционных программ.
В настоящее время идет интенсивная разработка основ генетики количественных признаков с привлечением математических методов. Несомненно, что эффективность этих методов может быть значительно повышена при расшифровке природы полигенов, определяющих количественные признаки растений. Дело в том, что морфологические особенности исходного селекционного материала не всегда позволяют прогнозировать свойства гибридных растений, поэтому целесообразно оценить наследуемость более простых слагаемых признака продуктивности растений. В этом плане представляет интерес поиск и фенотипическая идентификация генетических систем, обладающих сильным плейотропным эффектом на формирование признака продуктивности.
О существовании таких относительно немногочисленных генетических систем свидетельствует ряд экспериментальных данных. Так, например, показано, что явлению гибридной мощности сопутствует сравнительно небольшой оптимум гетерозиготности. Наиболее убедительным в этом отношении является эффект моногибридного гетерозиса. Известно также, что, несмотря на относительно небольшой вклад многих генов в формирование признаков высшего иерархического порядка, имеется ряд моногенно наследуемых признаков, существенно влияющих на габитус отдельных индивидуумов и на продуктивность популяции в целом. Полагают, что эти специфические гены локализованы в тесно сцепленных блоках полигенов и проявляют свой эффект только в составе определенных генотипов.
Совершенствование селекционных программ на основе использования принципов генетики количественных признаков позволит в дальнейшем более эффективно тестировать признаки продуктивности лесных древесных. Особенно перспективной в этом плане является разработка программы селекции по оптимизации фотосинтетических групп ценозов. О значимости этого направления исследований, в частности, свидетельствует уменьшение величины индекса облиственности у высокопродуктивных межвидовых гибридов тополей.
По-видимому, для всех категорий гетерозиса (индивидуальный, групповой, популяционный) лесным древесным растениям свойственна оптимальная гетерозиготность, обеспечивающая высокую приспособляемость к изменяющимся условиям среды. Наряду с этим в повышении гомеостатичности растений важную роль, играет, вероятно, оптимизация всех метаболических процессов, что, в свою очередь, способствует эффективному использованию ресурсов окружающей среды и экономному расходованию энергетических ресурсов в самом растении. Общепризнано, что гетерогенность или полиморфизм популяций является стратегией выживаемости, основой адаптации к изменяющимся условиям среды. Нами экспериментально установлено, что для гибридов с адаптивным гетерозисом гетерогенность структур проявляется на всех уровнях организации растений; на организменном (гетерогенность составляющих биомассы), тканевом (гетерогенность анатомических структур), клеточном (структурная гетерогенность хлоропластов), молекулярном (гетерогенность молекулярных форм ферментов). В связи с этим гетерогенность, видимо, следует считать универсальным механизмом адаптации.
Рассмотренные выше методы и принципы генетического анализа растений на потенциальную продуктивность позволяет в будущем вести селекционный процесс более планомерно и целенаправленно. Вместе с тем, уже в настоящее время на основе комплексного изучения генетических процессов у лесных древесных на разных уровнях организации их жизнедеятельности можно предложить программы интенсификации селекции лесных древесных.
Анализ закономерностей наследования различных признаков и их вклад в продуктивность гибридов выявил возможность раннего прогнозирования различных типов гетерозиса по листовым индексам, активности меристем, характеру строения мезофилла листа, по термоустойчивости, по изофермент-ному спектру и особенностям нуклеинового обмена. В селекции лесных древесных растений с длительным жизненным циклом использование методов раннего прогнозирования позволит значительно сократить селекционный процесс и проводить отбор в раннем возрасте.
Знание генетико-эволюционных характеристик исходного материала, изучение формообразовательного процесса в исходном материале и закономерностей формообразования при гибридизации и полиплоидии, выявление особенностей проявления различных типов гетерозиса, разработка методов ранней диагностики продуктивности, все это позволяет разрабатывать целенаправленные селекционные программы генетического улучшения лесных древесных растений и выбирать нужные направления и методы селекции того или другого вида с учетом целевого направления лесохозяйственного использования улучшенного селекционного материала.
Дата добавления: 2017-08-01; просмотров: 1104;