Образование временных связей пищевого условного рефлекса.
После первых сочетаний индифферентного раздражения с безусловным пищевым
рефлексом, изолированная проба этого раздражения вызывает только ориентировочную реакцию головы: животное поворачивает голову в сторону пищи. Это происходит| благодаря репродукции образа местонахождения пищи. Лишь в результате многократного сочетания условного сигнала с подкармливанием, образуются временные связи между воспринимающими элементами звукового, вкусового и интероцептивного анализаторов.
Интероцептивные раздражения вступают во временные связи при подкармливании потому, что интероцептивный анализатор активируется при каждой реакции какого-либо внутреннего органа и может вступать во временную связь со всякого рода совпадающими раздражениями. Благодаря этому наступившая прирожденная вегетативная реакция будет усиливаться и удлиняться условнорефлекторным путем подобно тому, как при рефлекторных движениях конечностей раздражение проприоцепторов поддерживает двигательную реакцию.
По новейшим электрофизиологическим исследованиям, интероцептивный анализатор у млекопитающих перекрывает кожный анализатор. Установлено, что при электрическом раздражении брыжеечного нерва или механическом раздражении брыжейки, медленные и быстрые электрические потенциалы возникают в сигмоидной извилине и в передней части эктосильвиевой извилины. В наиболее выраженной форме они наблюдаются в противоположном полушарии, в той части его, которая воспринимает импульсы со стороны кожных рецепторов туловища; в меньшей степени они наступают и в ипсилатеральном полушарии (Амасьян, 9). При раздражении блуждающего нерва усиление электрической активности наблюдалось в орбитальной области коры больших полушарий (Бейли и Бремер, 10).
Весьма возможно, что премоторная зона коры является дополнительным ассоциационным полем для интероцептивного анализатора, в основном лежащего в задней сигмоидной и в передней — эктосильвиевой извилинах.
Это вполне согласуется с результатами опытов Быкова и сотрудников (1947), которые показали, что удаление премоторной зоны лишь в некоторой степени меняет условнорефлекторную регуляцию внутренних органов. Особенно тщательно были установлены пределы коркового представительства внутренних органов Черниговским и сотр. (11), показавшими, что афферентное и эфферентное представительство вегетативных органов занимает моторную и премоторную зону коры, передние отделы лимбической системы, орбитальную кору и лишь незначительно заходит в теменную область. По-видимому, при образовании слюнного условного рефлекса анализатор, куда адресован условный раздражитель, вступает во временную связь не только с вкусовым анализатором, но и с интероцептивным. Надо считать, что секреция на условное раздражение не может наступить без участия этого анализатора. Тот факт, что условный слюнной рефлекс образуется после значительно большего числа сочетаний, чем оборонительный рефлекс, повидимому, объясняется тем, что интероцептивный анализатор обладает меньшей способностью к образованию временных связей, чем двигательный анализатор.
Итак, можно утверждать, что когда временные связи между анализатором, куда адресован сигнальный раздражитель, и двигательным или интероцептивным анализатором достигают определенной степени развития, они начинают возбуждать эфферентную систему пирамидных нейронов для производства внешней реакции.
Однако известно, что когда афферентная система двигательного или проприоцептивного анализатора связана хорошо развитыми нервными путями с определенной эфферентной системой и в силу этого возбудимость этих путей, а также афферентных клеток значительно повышена в результате каждодневной условнорефлекторной деятельности, то такая афферентно-эфферентная система может включиться в реакцию на новое сигнальное раздражение значительно скорее после нескольких сочетаний с ним безусловного раздражения. Так происходит потому, что эти эфферентные
|
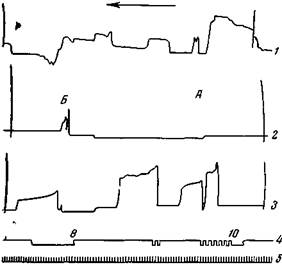
| Рис. 136. Рефлекторное сгибание передней правой лапы в ответ на электрическое раздражение задней лапы Предварительно рефлекс был образован на передней правой лапе. 1 — голова; 2 — задняя правая лапа; 3 — передняя правая лапа; 4 — отметка раздражения задней лапы; В — время, сек. В опыте А раздражение яапы слабое (10 см между индукционными катушками) и вызывает поднятие передней лапы. В опыте Б более сильное раздражение (8 см) вызывает сначала сгибание задней лапы, а потом поднятие передней |
|
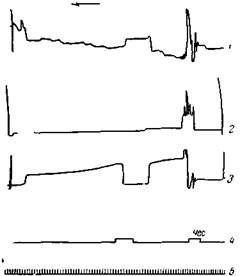
| Рис. 137. Опыты на той же собаке После образования условного рефлекса на передней лапе на звонок был образован новый рефлекс на правой задней на чесание левого бедра. Первая проба условного раздражения—чесание бедра—вызвало общее освободительное движение, а затем сгибание передней лапы; повторное чесание вызвало поднятие передней лапы с самого начала. 1 — голова; 2 — задняя правая лапа; 3 — передняя правая лапа; 4 — отметка раздражения лапы током и раздражения бедра чесалкой (чес.); 6 — время, сек. |
пирамидные клетки с повышенной возбудимостью легко активируются посторонними раздражениями. Это положение лучше всего можно иллюстрировать следующим опытом. Допустим, сначала был образован на звонок условный оборонительный рефлекс правой передней лапы. Когда этот рефлекс упрочился, приступили к образованию условного рефлекса правой задней лапы на чесание левого бедра. Первые пробы слабого электрического раздражения задней лапы обычно вызывают сгибание передней лапы; более сильные же — сначала сгибание раздражаемой задней, а затем поднятие передней (рис. 136). После нескольких сочетаний чесания бедра с электрическим раздражением задней лапы, чесание также начало вызывать с самого начала поднятие правой передней лапы или же общие освободительные движения (рис. 137). Лишь после многократного сочетания чесания левого бедра с электрическим раздражением правой задней лапы, чесание начало регулярно вызывать поднятие последней (Беритов, 12).
Дата добавления: 2017-01-13; просмотров: 661;