Судорожная электрическая активность.
Спонтанная электрическая активность коры мозга иногда принимает совершенно особый характер. В самой легкой форме это заключается в появлении более сильных двуфазных разрядов — быстрого колебания потенциала вначале и последующего медленного колебания в противоположном направлении. Обе фазы вместе могут продолжаться 0,05—0,2 сек. После каждого такого разряда спонтанная активность ослабевает или исчезает на разный срок, измеряемый десятыми долями секунды. Иногда они наступают на фоне дельта-волн (рис. 96). Эти ненормальные разряды могут наступить изолированно в одном участке, причем как спонтанно, так и в ответ на внешние раздражения. Так бывает, например, у животных при легком локальном отравлении коры большого мозга ацетилхолином или стрихнином (Дюссер де Баренн и сотр., 109; Беритов и сотр., 105). Если двуфазные разряды сопровождаются судорожным подергиванием соответствующей конечности, то их принято называть судорожными (Корнмюллер, 110). Но фактически они бывают в любом участке коры при достаточно высокой возбудимости, и когда они возникают изолированно в немоторной зоне, то не сопровождаются судорожным сокращением мышц.
При более или менее значительном локальном стрихнинном или ацетилхолиновом отравлении судорожные электрические разряды наступают длительными сериями. Высокая возбудимость отравленного участка является основным условием для наступления этих разрядов. Конечно, при этом условии они возникают как спонтанно, в результате самовозбуждения нервных кругов, так и под влиянием периферических импульсов. Но при сильных периферических раздражениях или при сильном раздражении самой коры судорожные разряды наступают и при нормальной или даже пониженной корковой возбудимости (Беритов, Гедеванишвили, Воробьев, 111). При сильном раздражении судорожные разряды появляются длительными сериями в правильном ритме от 4 до 20 в сек. Так бывает и спонтанно, если возбудимость была достаточно высокая (Корнмюллер, 110; Беритов и сотр., 8, 105), а также при эпилепсии и при наличии инородного тела в коре пли под корой при черепных ранениях (Беритов и сотр., 70, 99, 112).
Характерно, что когда очаг раздражения находится в коре, судорожные разряды являются локальными. Так бывает, например, при электрическом раздражении небольшого участка коры и при локальном отравлении небольшого участка коры такими активными веществами, как ацетилхолин и стрихнин. Было установлено, что при электрическом раздражении или локальном отравлении судорожные разряды не наблюдаются даже на расстоянии нескольких миллиметров от первичного возбуждаемого участка. Так. напри-
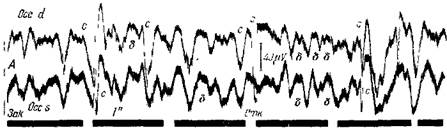
Рис. 97—I. Судорожные биотоки коры большого мозга у эпилептика
Биотоки отводятся от симметричных участков затылочной области. Один пластинчатый электрод (сетка) лежит на затылке, а другой (земля) — на мочке уха Глаза сначала закрыты, затем открываются. Верхняя запись от правого полушария дает более сильнье разряды, и притом более часто (с),чем от левого полушария Когда судорожный разряд справа наступает сравнительно сильно, тогда он проявляется и слева и притом совершенно синхронно Между судорожными разрядами сихронныс дельтаволны. Альфа-волны отсутствуют. Бета-волны сильно ослаблены. Такая же картина электрической активности наблюдалась у этого вш*лептика во всех других областях коры (Бсритов, Бакурадзе Дзидзишвили)
мер, при отравлении двигательной зоны передней ноги эти разряды не отводятся от двигательных зон задней ноги. Они отсутствуют даже в симметричных участках другого полушария (Дюссер де Баренн, 109; Беритов и сотр. 8, 15). Более того, в начальной стадии отравления ацетилхолином судорожные разряды могут возникать изолированно то в поверхностных, то в глубоких слоях коры отравленного участка (рис. 97—II).
В случае отравления двух участков коры мозга в каждом из них могут наступить судорожные разряды своим собственным ритмом, без заметного

Рис. 97 — II. Одиночные судорожные разряды коры большого мозга
Локальное отравление ацетилхолияо'я в обисти отаодящдх этзктрэцтз Нппьчый период отравления Верхняя кривая (Гл) — при биполярном отведении из нижних слоев коры мозга, нижняя кривая (Пов )—то же от поверхности мозга на том же уровне. Межполюзное расстояние в обоих случаях 5 «и В середине здпизи сильные судорожные разряды возникают раздечьно то в пуЗэктх, та в новзр^чозтлонс слояк (Беритов и Гедевани)
взаимодействия. Но во всех случаях, если судорожные разряды наступают в одном участке, в соседних участках того же полушария, а также в симметричном участке другого полушария, происходит торможение спонтанной электрической активности как быстрых, так и медленных колебаний.
Судорожные разряды, вызванные фарадическим раздражением коры, можно поддержать на долгое время раздражением определенных участков коры отдельными электрическими ударами и отдельными афферентными импульсами в ритме 1—4 в сек. (Розенблют и Кэннон, ИЗ). Это происходит благодаря повышению возбудимости в раздраженном участке. Участок первым долгом вовлекается в ответ на новое раздражение коры отдельными ударами. Было установлено, что передача судорожной деятельности от одного полушария к другому происходит по длинным комиссуральным нервным путям, а распространение их в пределах одного полушария — по коротким нервным связям: от раздраженных клеток — к соседним и так далее, все дальше и дальше от раздражаемого участка (Розенблют, Бонд и Кеннон, 114). Судорожные разряды в некоторых случаях наступают во всей коре, и притом совершенно одновременно, но интенсивность разрядов может быть в одном полушарии больше, чем в другом. Они могут наступить в одном полушарии при отсутствии их в другом. Это, например, может случиться при общем повышении возбудимости в коре, но при наличии очага раздражения вне коры в одной половине ствола мозга. Усиленные импульсы, исходящие из этого очага, достигают коры более или менее одновременно, но действие их будет сильнее в одном полушарии, чем в другом. Благодаря этому судорожные разряды будут проявляться в одном полушарии сильнее, чем в другом.
При локализации очага в неспецифической системе промежуточного мозга и ствола головного мозга, которая более или менее диффузно проецируется на кору, в ЭЭГ может регистрироваться диффузно синхронная судорожная активность (Пенфилд и Джаспер, 4; Гасто, 115).
Чрезвычайная характерность высокоамплитудных судорожных разрядов, которые намного превосходят нормальную фоновую активность, обусловила то, что электроэнцефалография стала важнейшим методом в диагностике эпилепсии. В зависимости от формы эпилепсии и локализации эпилептогенного очага описаны различные типы судорожных пароксизмальных разрядов. Но самой общей их чертой является то, что, как правило, эпилептические разряды являются выражением чрезмерно повышенной активности
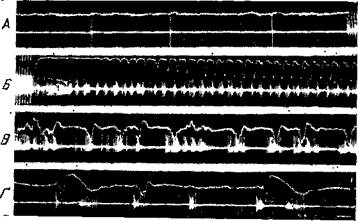
Рис. 98. Эпилептическая активность, вызванная электрическим раздражением коры
Верхняя кривая — электрокортикограмма, нижняя кривая — нейронная активность, зарегистрированная внеклеточным микрсэлектродом. А — перед электрическим раздражением коры и в начале раздражения. Б, В я Г — после электрического раздражения (начало и конец раздражения видны по артефактам). Между Б, В, и. Г перерывы в несколько сек. Калибровки: горизонтальная линия—100 мсеп, вертикальная— 200 мкв для электрокортикограммы (Окуджава)
нейронов (Моруцци, 116). Оказалось, что при максимальном развитии эпилептической активности в очаге ее около 95% нейронов данного участка приходят в состояние повышенной активности (Окуджава, 117).
Ясно, что понять природу эпилептической активности возможно только при учете тех изменений, которые происходят на уровне отдельных нейронов. Выявить эти изменения удается с помощью микроэлектродной регистрации деятельности одиночных нервных клеток в эпилептогенном очаге. На рис. 98 приводится одновременно с электрокортикографической кривой внеклеточная запись одного нейрона сенсомоторной области коры в норме и после развития эпилептической активности в результате локального ритмического электрического раздражения (Окуджава, 118). Видно, что по прекращении электрического раздражения одновременно с судорожным разрядом последействия, отводимым с поверхности коры, нейрон из среднего слоя дает высокочастотный разряд пиковых потенциалов, причем частота намного превосходит наблюдаемую в норме частоту разрядов данного нейрона. Вместе с тем, отмечается резкое уменьшение амплитуды пиковых потенциалов. Как видно на рис. 98, Б, после прекращения электрического раздражения продолжается сплошной высокочастотный разряд нейрона со значительно сниженной амплитудой пиковых потенциалов. Кроме того, в отличие от разряда нейрона в нормальных условиях, наблюдается значительная вариабильность амплитуды пиковых потенциалов, с их уменьшением порой до полного исчезновения. При этом колебания амплитуды коррелируют с пароксизмальными волнами на электрокортикограмме (рис. 98, Б, В, Г).
Такие изменения нейронных разрядов становятся понятными при учете сведений, полученных с помощью внутриклеточной регистрации деятельности нейрона. На рис. 99 приведена внутриклеточная запись нейрона при аналогичной постановке эксперимента. Видно, что при развитии эпилептической активности наблюдается продолжительная деполяризация мембраны нейрона, вызывающая высокочастотный разряд пиковых потенциалов. На фоне такой сплошной деполяризации отмечаются отдельные, большой амплитуды и продолжительности' медленные колебания мембранного потенциала — пароксизмальные деполяризационные сдвиги (рис. 99, Б, Б, Г). Каждый такой деполяризационный сдвиг вызывает учащение разрядов вплоть до полной инактивации генерации пиков (рис. 99, Б). Амплитуда деполяризационных сдвигов не всегда максимальная. Например, на рис. 99, Б видно
четыре таких колебания все нарастающей амплитуды. Эти деполяризационные сдвиги коррелируют с пароксизмальными волнами электрокортикограммы. При продолжающейся эпилептической активности мембрана все больше и больше деполяризуется, деполяризационные сдвиги становятся все более продолжительными, механизм генерации пиковых разрядов полностью инактивируется, вместо полноценных пиков временами отмечаются незначительные осцилляции. На этом деполяризованном фоне в электрокортикограмме могут отмечаться деполяризационные сдвиги, не связанные с пиковыми разрядами.
Как видно из приведенных данных Окуджавы, эпилептическая активность нейронов характеризуется чрезмерно высокой частотой разрядов и вариабельностью амплитуды пиковых потенциалов. Автор полагает, что в основе такого явления лежат значительные деполяризационные изменения мембранного потенциала нейронов. На основе микрофизиологического анализа было сделано заключение, что под воздействием эпилептогенного фактора, которым может быть любое достаточно сильное раздражение, вызывается длительное возбуждение большого числа нейронных элементов. В результате этого в синаптических окончаниях происходят длительно сохраняющиеся потенциационные изменения. После этого передача импульсов по этим синаптическим окончаниям вызывает не обычные деполяризационные постсинаптические потенциалы, которые регистрируются при нормальной спонтанной активности, а вышеотмеченные пароксизмальные деполяризационные сдвиги. В пользу такой постсинаптической природы говорит, с одной стороны, градуальный характер этих колебаний (рис. 98, Б), а с другой стороны — то, что они могут возникать и тогда, когда механизм генерирования пиковых потенциалов полностью инактивирован (рис. 99, Г).
Так как с эпилептическими волнами электрокортикограммы на клеточном уровне в средних слоях коры коррелируют отмеченные пароксизмальные деполяризационные сдвиги мембранного потенциала, нужно думать, что эпилептические волны в электрокортикограмме являются суммарным отражением таких сдвигов (Окуджава, 118).
Но весьма возможно, и это было бы намного вероятнее, что здесь дело идет не об особом патологическом изменении природы клеточной постсинаптической мембраны, а о необычайно большом увеличении количества акти-
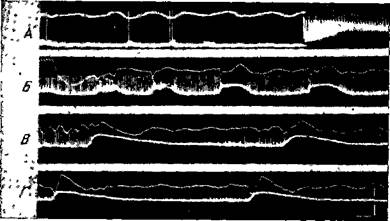
Рис. 99. Активность нейрона, зарегистрированная при помощи внутриклеточного микроэлектрода
Верхняя кривая — электрокортикограмма, нижняя — активность нейрона А — перед раздражением коры и начало электрического раздражения. В — конец раздражения и непосредственно после него. Между Б, В и Г перерывы в несколько сек Калибровки: горизонтальная линия— 100 мсек, нижняя вертикальная — 20 мв для микроэлектрода и верхняя вертикальная — 400 мкв для макроэлектрода (Окуджава)
•вированных дендритных синапсов на пирамидных нейронах. Вызываемые ими суммарные длительные электротонические токи заходят в пирамидные клетки, а выходя из них в синаптических участках, вызывают при некоторой малой амплитуде длительное возбуждение клеток спиковыми потенциалами, а при большой амплитуде — их катодическую депрессию. Пиковые потенциалы, отводимые из сред них слоев коры, часто сопровождаются медленными сдвигами потенциала на поверхности коры мозга. Это в свою очередь должно быть объясняется активирующим действием возбужденных возвратных коллатералей аксона пирамидных клеток на дендриты поверхностных слоев коры.
Происхождение судорожных разрядов объясняется усиленным возбуждением большого комплекса нервных кругов в раздраженном или отравленном участке, а затем и в соседних участках. Первая фаза каждого разряда зависит от возбуждения первично возбужденных нервных кругов (отравленного участка в случае самовозбуждения, раздраженного участка в случае раздражения), а вторая фаза — в противоположном направлении: от возбуждения ближайших нервных кругов в соседних участках. Усиленная иррадиация импульсов возбуждения из нервных кругов будет производить усиленное активирование дендритной массы поверхности коры как в первичном возбужденном, так и в ближайших участках. Потому активирование дендритов в последних участках будет начинаться и кончаться позднее, чем в первично возбужденном: сообразно при отведении первично возбужденного участка дендритный ток сначала должен протекать в одном направлении, а потом в другом. Кроме того, этот дендритный ток, согласно дендритной гипотезе торможения, должен производить наблюдаемое при этом угнетение спонтанной электрической активности как в судорожно-возбужденных участках коры, так и в соседних участках.
Дата добавления: 2017-01-13; просмотров: 1406;