ГЛАВА VII. ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА И ЕЕ ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ
Общее понятие.
Кора большого мозга, подобно другим отделам головного мозга, непрерывно продуцирует электричество, источником которого является активное состояние нервных и, возможно, глиальных элементов. На животных корковая электрическая активность изучается главным образом путем отведения биотоков непосредственно от обнаженного мозга. На людях ее изучают главным образом путем отведения от кожной поверхности черепа, куда биотоки коры распространяются физически через оболочки мозга, кости черепа и кожу. Потенциалы, генерируемые корой, измеряются при отведении от поверхности мозга обычно долями милливольта, а при отведении от кожи черепа — десятками микровольт (миллионных долей вольта).
Биотоки, отводимые от поверхности данного участка коры, отражают физиологические процессы, протекающие в отводимом участке, а не в остальной коре или подкорковых структурах. Биотоки соседних участков могут отводиться вследствие чисто физического распространения только в том случае, когда они очень большой интенсивности, притом эти биотоки будут отводиться чрезвычайно ослабленными (Беритов, Брегадзе и Цкипуридзе, 1; Ройтбак, 2).
При отведении от кожи черепа корковые биотоки сильно ослаблены, вследствие побочного замыкания их через тканевую жидкость (Беритов, 3). В последнее время специальными опытами и математическими расчетами точно установлено шунтирующее действие окружающих поверхность коры тканей, именно церебро-спинальной жидкости, мышц, мягкой и твердой мозговых оболочек. Эти факторы приводят к ослаблению частых колебаний в 20 раз, а медленных в 5 раз (Пенфилд и Джаспер, 4).
Электрическая активность коры, как и других отделов мозга, состоит из длительных колебаний чрезвычайно изменчивой продолжительности. Такие нерегулярные медленные потенциалы свойственны всем отделам мозга (рис. 82). Наряду с ними в коре нормальных животных и человека всегда наступают сравнительно быстрые колебания потенциала также меняющейся продолжительности; по амплитуде они обычно значительно ниже медленных. Быстрые колебания потенциала наступают обычно на фоне медленных. Быстрые потенциалы тоже наступают во всех отделах мозга. Таким образом, медленные нерегулярные потенциалы и быстрые на фоне их характеризуют «спонтанную» деятельность всех частей коры больших полушарий при ее нормальном функционировании, а потому можно эту электрическую активность считать основной электрической активностью коры (рис. 83).
У человека в определенных отделах коры и при определенных условиях временами устанавливается правильный ритм медленных колебаний с продолжительностью около 0,1 сек. Этот ритм впервые был открыт в 1929 году Г. Бергером (5), потому его называют бергеровским ритмом, но чаще его называют альфа-ритмом, а составляющие его волны — альфа-волнами. Альфа
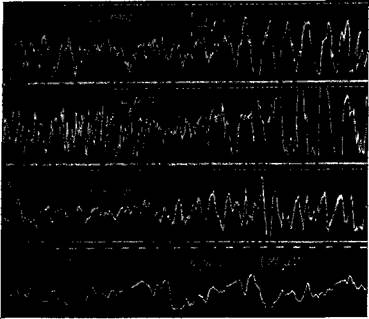
Рис.82. Электрическая активность коры большого чозга кошки
Катодный осциллограф Биполярное отведение обнаженной коры с межполюсным расстоянием 3 мм: А — от лобной доли, В — от' теменной, С — от затылочной (во всех случаях сначала дается период,сравнительного зтгишья, а затем период усиленной деятельности),!) — от теменной доли при более быстром] вр-нцении фотографического аппарата (Беритов, Бр^адзе и Цкипуридзе)
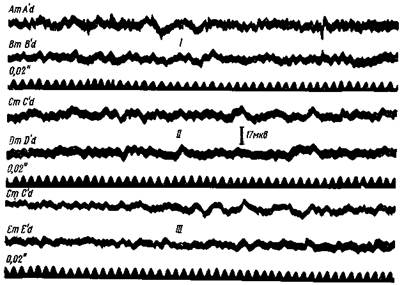
Рис. 83. Основная электрическая активность коры большого мозга человека
Регистрация биотоков шлейфным осциллографом Поперечное биполярное отведение от кожной поверхности черепа с правой стороны, как показано на рис 85 Межполюсное расстояние 5—6 см АтА' — биотоки от затылочной области, ВтВ' — оттененной области; CmC' — от задней границы постцентральной извилины, DmD' — от передней границы прецентральной извилины, ЕтЕ' — от лобной об части Все осциллограммы засняты при открытых глазах на свету
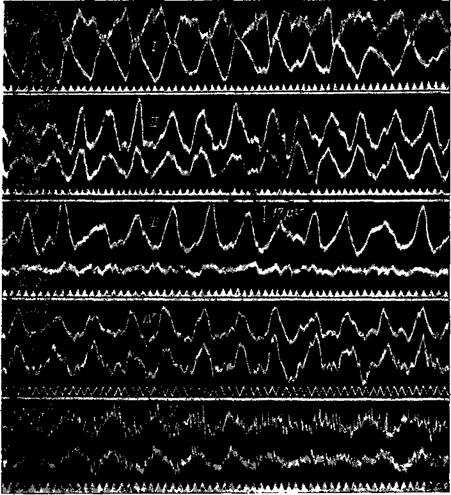
Рис. 84. Альфа-ритм коры большого мозга человека
Регистрация шлейфным осциллографом Отведение биотоков продольное, биполярное, с правой стороны Положение электродов, как на рис. 85 АВ — биотоки от затылочной области; ВС — от теменной; CD — от центральной области (постцентральной и прецентральной извилин); DE — от задней половины лобной области, GH — от височной области Все осциллограммы — при закрытых глазах. Осциллограммы I—IV от того же субъекта, что на рис. 83, а V — от другого. Везде хорошо выраженные альфа-волны, кроме лобной доли, где они совершенно отсутствуют
волны наступают у человека, как правило, в затылочных и теменных долях и обычно отсутствуют в лобных долях; в височных и центральных долях они наступают не с такой интенсивностью, как в затылочных и теменных, и не всегда (рис. 84). Обычно отводимые участки черепа и соответствующие участки коры большого мозга представлены на рис. 85 и 86.
В настоящее время в записях от черепа человека (в электроэнцефалограммах — ЭЭГ) колебания электрического потенциала классифицируются на основании их частоты, определяемой автоматическим устройством — частотным анализатором, следующим образом: от менее 1 до 3,5 в сек. (т. е. продолжительностью от более 1 сек. до 0,03 сек.) — дельта-волны; от 4 до 7 в сек. — тета-волны; от 8 до 13 в сек.— альфа-волны; от 14 до 100 и более в сек — бета-волны (Уолтер, 6).
Все перечисленные типы электрических потенциалов имеют особое происхождение и связаны с определенным состоянием организма. Так, бета-волны у животных и человека доминируют при деятельном состоянии — при
|
интенсивной физической и умственной работе, при осуществлении ориентировочных и увловных рефлексов. Альфа-волны возникают у человека при исключении зрительных раздражений: при выключении света или при закрытии глаз, в состоянии относительного покоя (рис. 87). Дельта-волны характеризуют у взрослого нормального человека и у животных состояние естественного сна. Предполагается связь тета-волн с некоторыми эмоциональными состояниями (Уолтер, 6), но они регистрируются в центральной области при статической мышечной работе (Ройтбак, Дедабришвили и Гоциридзе, 7).
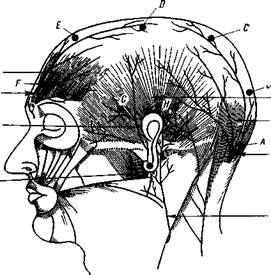
| Рис. 85. Расположение мышц и нервов на голове и электродов для продольного биполярного отведения Точки обозначают местоположение отводящих электродов Они расположены на расстоянии 3 см от медиальной линии Точка А — на задней границе затылочной области на Imea nuchae, В — на передней границе затылочной области, С — на передней границе теменной области, D — на передней границе прецентрал ьной извилины; Е — на середине лобной области, F— на передней границе лобной области, Н и G — в виоочн ой области Между соседними точками — 5—6 см При наложении электродов на точки А и В отводится ЭЗГ затылочной области, на В и С — теменной, С и D — центральной (постцентральная и прецентральная извилины), D и Е — задней половины лобной области, на Е и F — передней половины лобной области, Н и G — височной области |
|
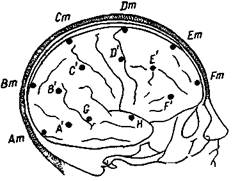
| Рис. 86. Расположение электродов на голове при поперечном отведении и отношение их к долям большого мозга Am—area stnata, A'— на границе ar praeoccipit" ahsuar occipito-temporahs, Вт — аг praeoccipitalis, В' — на границе ar angularis и ar. temporalis sup , Cm— на границе аг parietalis и аг praepanetalis, С' — на границе аг. аг. supramargmalis, temper sup и temp transversa post ; Dm — ar praecentrahs gigantopyramidahs, D' — нижний край ar postcentralis, Em — на границе ar. front intermedia и аг frontalis gran , E' — ar opercularis, Fm — praefrontalis, F' — ar tnangulans Между соответствующими электродами около 5 см Кроме того, два электрода G и Н на височной области |
Когда функциональное состояние коры улучшается (в условиях острого опыта) и возбудимость ее нервных элементов повышается, спонтанная электрическая активность усиливается: повышается амплитуда медленных и быстрых колебаний потенциала. То же наблюдав!ся при повышении возбудимости от любой причины, например при отравлении коры эзерином (Беритов, Брегадзе и Цкипуридзе, 8). Когда же функциональное состояние коры ухудшается и корковая возбудимость падает, спонтанная электрическая активность ослабевает: уменьшается амплитуда медленных и быстрых колебаний основной активности. Это бывает, например, при охлаждении коры, при отравлении ее кокаином (Беритов и сотр., 8), при анемии мозга, вызванной перерезкой сосудов (Правдич-Неминский, 9; Андреев и Шпильберг, 10). В коре больших полушарий кошки после остановки кровообращения сначала исчезают быстрые колебания,а потом, через 20 сек., исчезает вся электрическая активность, причем в коре она исчезает раньше, чем в других отделах головного мозга (Джаспер, 11); то же происходит при искусственной задержке дыхания.
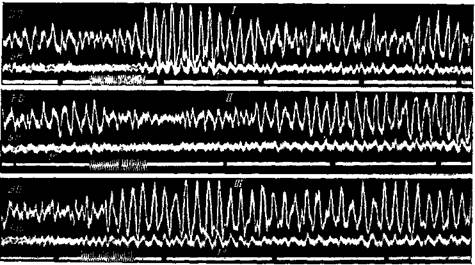
Рис. 87. Влияние закрытия или затемнения глаз на биотоки коры большого мозга человека
Шлейфный осциллограф. Отведение от теменной и ^центральной областей (BD) и от височной области (ng) I — сначала глаза открыты и в кабине светло, затем глаза по приказу закрываются. Альфа-волны усилились в первую те секунду. II — вначале глаза открыты и в кабине светло, затем свет тушится, в кабине абсолютная темнота, но глаза остаются открытыми. Альфа-волны усиливаются приблизительно через 2 сек. III — сначала темно, глаза открыты. Через несколько секунд от начала затемнения, когда альфа-волны ослабли, глаза по приказу закрываются. В ту же секунду вновь возникают усиленные альфа-волны. Время закрытия или затемнения глаз отмечается на линии отметки времени (Беритов и Воробьев)
Основная электрическая активность коры не зависит от импульсации со стороны подкорковых образований. Она по существу не меняется, если кору отделить от подкорковых образований путем перерезки всех восходящих и нисходящих путей. Она также включает альфа- и бета-волны и также меняется при функциональных изменениях коры, как нормальная (Хананашвили, 126).
Основная электрическая активность коры исчезает в небольшом участке коры, если он изолирован не только от подкорковых образований, но и от остальной коры. Но когда такая полоска коры имеет связь в виде мостика с остальной корой, тогда она дает основную электрическую активность (Берне, 75). Эти факты ясно указывают, что основная электрическая активность целиком является функцией коры большого мозга. При не нормальных условиях организма и при заболеваниях регистрируются патологические потенциалы, например, судорожные разряды при эпилепсии. Клиническая электроэнцефалография развилась в специальную отрасль исследований и метод этот играет важнейшую роль в диагностике и прогнозировании заболеваний мозга (Пенфилд и Джаспер, 4; Чугунов, 12).
Дата добавления: 2017-01-13; просмотров: 1504;