Влияние ретикулярной формации на кору больших полушарий.
Впервые в 1949 г. Моруцци и Мэгун (1) показали, что при электрическом раздражении ретикулярной формации протекающая до того в коре большого мозга электрическая активность в виде медленных колебаний, характерная для дремотного состояния животного, затухала и заменялась быстрыми потенциалами малой амплитуды (эффект десинхронизации ЭЭГ, которая на ненаркотизированном препарате длилась известное время и после прекращения раздражения) (см. также Нарикашвили, 2). Таким образом наблюдалась картина электрической активности, характерная для пробуждения или бодрствующего состояния животного (Рейнбергер и Джаспер, 3; Морисон, 4; Цкипуридзе, 5) (рис. 101). Если такой же опыт провести на животных в хронических условиях с вживленными в мозг электродами, то в ответ на тетаническое раздражение ретикулярной формации дремлющее или спящее животное просыпается (Сегундо, Арана, Френч, 6). Значит, пробуждение животного и ассоциированная с ним электрическая активация коры, что часто называют также «реакцией пробуждения», развивается благодаря действию восходящих ретикулярных импульсов. Несмотря на довольно подробное изучение особенностей восходящей активирующей системы, до сих пор еще не установлены те пути, по которым ретикулярные импульсы достигают коры больших полушарий. Предполагали путь через неспецифические (медиальные) таламические ядра, однако окончательно восходящие к коре пути из этих ядер еще не установлены. Возможно ретикулярные импульсы распространяются к коре, проходя через вентральное переднее ядро и передний полюс ретикулярного ядра таламуса, которые, судя по их ретроградной дегенерации после декортикации, имеют прямые связи с корой. Однако, это, видимо, не единственный путь, по которому ретикулярные импульсы достигают коры. Описаны пути, идущие как по субталамусу, так и через палеокортекс и полосатое тело.
Если бодрствующее состояние животного поддерживается восходящими ретикулярными импульсами, то при прекращении или резком ограничении поступления их в кору у животного в отводимой ЭЭГ должна развиться картина, характерная для сна, и оно должно заснуть. Так получается и на самом деле: при перерезке ствола головного мозга на уровне четверохолмия, лишающей кору влияний главной массы ретикулярной формации (рис. 102), в коре развивается синхронизированная активность в виде медленных волн и периодических вспышек веретен (Линдсли, Мэгун, 7,8,9; Нарикашвили и Мониава, 10), и животное впадает в глубокое коматозное состояние. Явление это было известно и раньше (Бремер, 11), но тогда оно получало другое объяснение.
Эти опыты с повреждением ретикулярной формации говорят о том, что ретикулярные импульсы в естественных условиях, так же, как и в случае нисходящего влияния, действуют на кору постоянно, тонически, периодически то усиливаясь, то ослабевая. Этим самым уровень бодрствующего состояния животного то повышается, то понижается. На основании всех этих данных и сложилось представление о ретикулярной восходящей активирующей системе (12) (рис. 100).
Исходя из этих фактов, естественный сон начали рассматривать как результат снижения уровня активности ретикулярной формации, вследствие чего снижался и тонус коры больших полушарий (13). Снижение же активности ретикулярной формации наступает главным образом благодаря ограничению периферических раздражителей, обычно поддерживающих активность последней на высоком уровне. Механизм наркоза получил также новое объяснение. Наркотические вещества, особенно барбитураты, оказывается

|
Рис.100. Афферентные связи ретикулярной системы
Афферентные волокна седалищного нерва от кожно-мышечных рецепторов (п. J) и от слухового нерва (VIII). Ретикулярная формация представлена черным цветом, ядра среднего и промежуточного мозга — точками. Афферентные слуховые прямые пути (LL) и афферентные ко «но-мышечныв прямые пути (Lm) отдают коллатерали к ретикулярной формации. Cm — центральное ядро; VP—задневентральное ядро таламуса; Gm — среднее коленчатое тело; DM — дорсомедиальное ядро; CS — сенсомоторная кора; СА — слуховая кора (по Френчу и сотр.)
в первую очередь действуют на ретикулярную формацию, снижая ее активность и ее восходящее влияние на кору больших полушарий. Это было показано в прямых опытах с одновременной регистрацией электрических ответов в ретикулярной формации и проходящих рядом афферентных путях. Оказалось, что при постепенном увеличении концентрации наркотика раньше всего ослабевают и пропадают ответы в ретикулярной формации. Большую чувствительность ретикулярной формации к наркотикам и вообще к
депрессантам связывают с мультинейронным строением восходящих путей, а так как те или другие фармакологические агенты или продукты обмена веществ оказывают свое действие путем влияния на синаптические образования, то отсюда ясно, почему наркотики должны снижать активность, прежде всего, ретикулярных образований.
За последнее время, главным образом работами, выполненными в лаборатории Моруцци, было показано, что наряду с восходящим активирующим десинхронизирующим влиянием из ретикулярной формации в кору поступает также синхронизирующее влияние. По представлению Моруцци и его сотрудников, верхняя часть ретикулярной формации ствола головного мозга, в частности ретикулярные образования среднего мозга и ростральной половины моста (верхний ствол), обладают десинхронизирующим влиянием на корковые нейроны, тогда как ретикулярная формация каудальной половины моста и продолговатого мозга (нижний ствол) влияют синхронизирующе. Такое заключение было вынесено прежде всего на основании опытов с перерезками мозга на различном уровне ствола головного мозга (Моруцци и сотр., 14; Моруцци, 15). Только после перерезки мозга по средней линии моста, сейчас же впереди от ядра тройничного нерва (средне-мостовой претригеминальный препарат) наблюдалась длительная десинхронизированная (активированная) картина ЭЭГ (рис. 103). По мнению авторов, это обусловлено тем, что при такой перерезке отключается от коры синхронизирующее влияние нижнего ствола, благодаря чему десинхронизирующее влияние верхнего
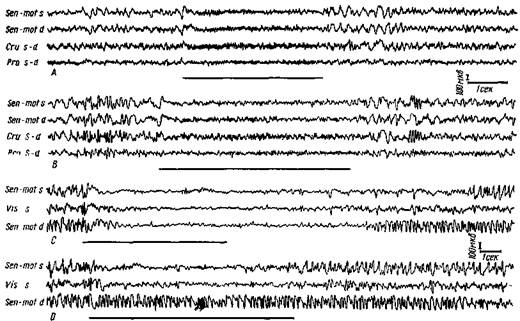
Рис. 101. Влияние раздражения ретикулярной формации ствола головного мозга иа электрическую активность коры больших полушарии кошки
А и В — изолированный энцефалический препарат кошки (изолированная голова). Для создания необходимого фона медленной активности введено малое количество хлоралозы (7 мг/кг). В каждом опыте сверху вниз отводятся потенциалы левой и правой сенсомоторной коры, левой крестовидной извилины и левой прореальной извилины. При раздражении бульварной ретикулярной формации (горизонтальная линия под каждой осциллограммой) левой половины мозга (1,5 в, 300 в сек.) хорошо видна замена высокоамплитудных медленных волн низкоамплитудной частой активностью в обоих полушариях. С —кошка с интактной нервной системой, хлоралозный наркоз (50 мг/кг). Отводятся сверху вниз: левая сенсомоторная кора, левая зрительная и правая сенсомоторная кора. При раздражении левой бульбо-ретикулярной формации (3 в, 300 в сек) угнетаются медленные волны билатерально, угнетение быстрее наступает и позже проходит в гомолатеральном полушарии. Хорошо видно, что при глубоком наркозе низковольтная активность не возникает.
D — те же условия пыта, как в С, но частота раздражения ретикулярной формации уменьшена до 100 в сек. Эффект ограничивается гомолатеральным полушарием с прекращением ретикулярного раздражения (Моруцци и Мэгун)
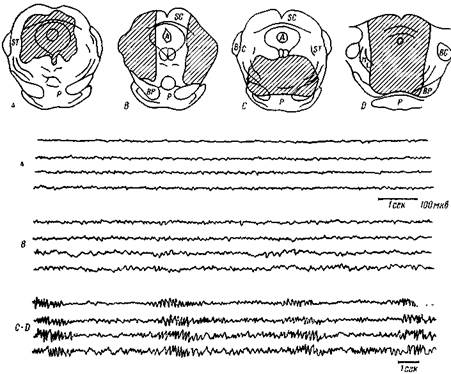
Рис. 102. Изменения электрической активности коры при повреждении серого вещества мозга
Наверху — схемы поперечных разрезов через средний мозг кошки с указанием повреждения (заштриховано) центрального серого вещества (А), латеральных проводящих путей (В) и покрышки среднего мозга (С). Они произведены на изолированном энцефалическом препарате кошки. Соответствующие изменения электрической активности коры приведены в нижней части рисунка. Во всех осциллограммах регистрируются сверху вниз потенциалы: коры передней части полушария слева и справа, коры задней части полушария слева и справа.
А — электрическая активность после повреждения центрального серого вещества вокруг сильвиева водопровода — электроэнцефалограмма (ЭЭГ) сохраняет «активированный» (десинхронизированный вид, медленная активность не развилась.
В — электрическая активность записана через 6 час. после повреждения восходящих путей (перереака В) и через 3 часа после перерезки зрительных нервов и разрушения обонятельных луковиц с обеих сторон. ЭЭГ слегка синхронизирована (по сравнению с А), но нет вспышек веретен, характерных для дремоты и сна. С — D — в ЭЭГ появляются частые вспышки веретен вскоре после изолированного повреждения вентральной части покрышки среднего мозга (перерезка С). Через некоторое время частота вспышек уменьшается, видимо, по той причине, что более дорзальная часть покрышки не повреждена. После широкого повреждения покрышки (перерезка D) возникали такого же характера вспышки веретен» которые сохранялись в течение всего времени наблюдения (по Линдсли, Боуден и Мэгун)
ствола проявляется в полной силе. В последующее время эти данные были подтверждены другими опытами. В одном из таких опытов соответствующими перевязками сосудов, снабжающих головной мозг, удалось добиться такого положения, что верхний ствол снабжался кровью только через сонную артерию, а нижний — через позвоночную артерию. Когда барбитурат, который, как известно, легко угнетает деятельность ретикулярных образований, вводился через сонную артерию, то из-за снижения активности ретикулярной формации верхнего ствола в усиленной форме проявилось синхронизирующее влияние из нижнего ствола, в результате чего в коре появилась синхронизированная активность. В случае введения барбитурата через позвоночную артерию, наоборот, наблюдали картину десинхронизации ЭЭГ. В последнем случае барбитурат снижал активность ретикулярной формации нижнего ствола, освободив этим самым от противоположного влияния ретикулярную формацию верхнего ствола, которая и проявила свое десинхронизирующее влияние.

Рис. 103. Изменения электрической активности коры больших полушарий после средпемостовой (А) и переднемостовой (В) перерезок
В нижней части рисунка показана ЭЭГ. Потенциалы отводятся от правой и левой лобных долей. Хорошо видно состояние десинхронизации ЭЭГ после среднемостовой перерезки (сейчас же впереди от мостового сенсорного ядра тройничного нерва — У) и возникновение синхронизированной (в виде вспыщек веретен) после переднемостовой^перерезки (В), выключающий влияние десинхронизирующего механизма на кору С. г. —нижнее двухолмие; 01. г. — верхняя олива; р. — ножки моста; N. с.—наружное клинообразное ядро; N. I. I. — ядро латеральной петли; Тг. sp. V — спинальный тракт тройничного нерва (Моруцци и сотр.)
По представлениям Моруцци, в естественных условиях разные периферические раздражения, в зависимости от того, применяются ли они первый раз или многократно, могут преимущественно возбудить то одну, то другую часть ретикулярной формации. При первых применениях раздражения активируется преимущественно десинхронизирующий ЭЭГ механизм. Если же раздражение повторяется монотонно большое количество раз, то оно все более и более активирует синхронизирующий механизм. Это было показано в опытах с длительным повторением (в связи с изучением явления «привыкания» ответов) световых раздражений, когда на самом деле ЭЭГ делается все более и более синхронизированной (появляются нерегулярные медленные волны и «веретена»). Так как такая картина ЭЭГ соответствует дремотному состоянию, то есть основание считать, что сон наступает не только в результате ослабления восходящего активирующего (десинхронизирующего) влияния, но и от постепенного усиления действия синхронизирующего механизма (см. также Дел и сотр., 16).
Но еще раньше нашим сотрудником Ройтбаком в опытах на кошках с вживленными электродами было показано, что при повторных применениях звуковых и электрокожных стимулов после первичных ответов в соответствующих воспринимающих областях коры начинают возникать медленные отрицательные потенциалы большой амплитуды; затем в остальной коре начинают доминировать медленные колебания; это сопровождается угнетением быстрых колебаний, а поведенчески — угасанием ориентировочного рефлекса на данное раздражение и развитием сонливости. По представлениям Ройтбака, «синхронизация» ЭЭГ и явления угнетения корковой деятель-
ности при этом обусловлены вовлечением в деятельность ретикулярной формации таламуса: при монотонных раздражениях возбуждение «проторяет» путь через многонейронные цепочки этого образования и посредством неспецифических афферентов достигает коры, активируя преимущественно дендритное сплетение верхних слоев. Предполагается, что в основе угасания корковых рефлексов лежит деятельность этого механизма. По Моруцци, таким механизмом является синхронизирующий механизм в нижней стволовой части ретикулярной формации (15).
Дата добавления: 2017-01-13; просмотров: 2800;