Торможение пирамидных нейронов.
Каждое возбужденное волоконце, прилегающее к дендриту, возбуждает через боковые синапсы массу шипиков на его стволе, а своим терминальным синапсом — окончания дендритов в первом слое. Причем активация этих синапсов происходит не в один момент, а последовательно. Поэтому в дендрите возникает суммарный, медленно нарастающий ток, который, распространяясь по дендриту электротонически, достигает и сомы клетки. При действии ряда нервных импульсов дендритные токи, сливаясь, образуют сплошной, значительной величины ток, который длится все время, пока происходит синаптическая бомбардировка дендритов или, точнее, постсинаптическая активация дендритов (рис. 36).
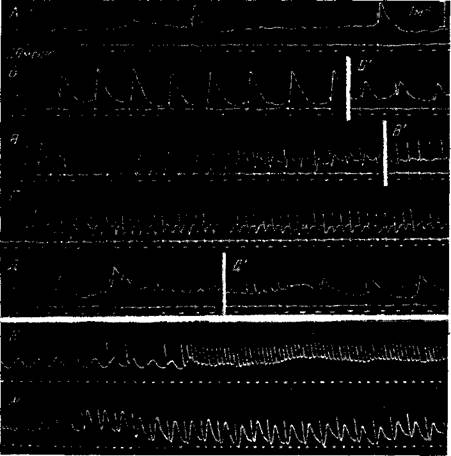
Рис. 35. Отрицательные потенциалы, вызываемые электрическим раздражением поверхности коры, при разной частоте раздражения
Отводящий и раздражающие электроды расположены на поверхности с>пр,юильвиевой извилины коры кошки, отводящий электрод находится на расстоянии !,"> мм от раздражающих. Интенсивностьраздражения 30 е (порог 8 е). Нембутал. А — частота раздражения около 3 в сек, Б — 16 в сек , Б' — эффекты после 1 мин. раздражения, В — частота раздражения около 30 в сек., В' — через 30 сек. раздражения; Г—частота раздражения 100 в сек. Д—отводящий электрод отодвинут на расстояние 2,Ьмм от раздражающих; частота рачдраженип с 15 в сек. мгновенно переключается на 100 в сек, Д1—после 1 мин.раздражения с частотой 100 в сек. происходит переключение на частоту 1о в сек , Е—после 3 сек. раздражения при частоте 25 в сек тетанизация переводится на частоту 12Г> в сек., Ж — после 15 сек тетанизапии при частоте 125 сек частота раздражения переводится обр itho на 25 в сек. (Ройтбак)
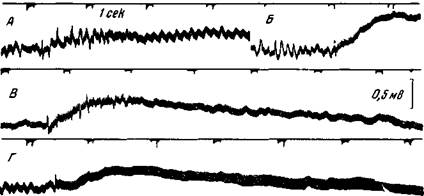
Рис. 36. Длительные потенциалы, возникающие при ллектрическом раздражении поверхности коры Кошка под глубоким нембуталовым наркозом На поверхности супрасильвиевой извилины установлены раздражающие электроды и на расстоянии 3 и 10 мм от них — отводящие электроды. Усилитель с большой постоянной времени Продолжительность раздражающих стимулов — 0,5 мсек А — частота раздражения 9 в сек , Б — частота раздражения 50 в сек , В — эффект повторного раздражения через 10 сек , во время тормозящего последействия. Интенсивность раздражения А — В — 10 в, Г — при частоте 100 в сек , 5 в Отклонение вверх означает отрицательность ближайшего к пункту раздражения отводящего электрода, при всех этих раздражениях коры «спонтанная» электрическая активность исчезает (Беритов и Ройтбак)
Дендритные токи, достигнув клетки и затем распространяясь по клетке, должны выходить из нее, чтобы вернуться к возбужденным участкам дендритов. Выходить же они должны в синаптических участках, где клеточные мембраны обладают большой электропроводностью, благодаря постоянному действию активных веществ, выделяемых синаптическими пузырьками. После выхода из клетки они должны проникнуть в нервные окончания. Здесь они вызывают, с одной стороны, анэлектротоническую блокаду пресинаптической мембраны, а с другой— возможно, отвлекают синаптические пузырьки от этой мембраны в противоположную сторону.
На основании электронномикроскопического исследования известно, что пузырьки собираются у пресинаптических мембран только во время возбуждения. Без этого они распространяются по всей клетке. Поэтому можно предположить, что при дендритном анэлектротоническом токе пузырьки передвигаются в сторону пресинаптического волоконца. Их химическое воздействие вместе с катэлектротоническим действием выходящего из волоконца дендритного тока может привести к значительной деполяризации пресинаптического волокна. Деполяризация пресинаптических волокон во время торможения была отмечена многими авторами (Экклс, 42). Понятно, что как деполяризация пресинаптического волоконца, так и анэлектротоническая блокада пресинаптической мембраны, создают препятствие для подходящих к телу клетки пресинаптических импульсов возбуждения (Бериташвили, 20, 43).
Это явление деполяризации пресинаптического волоконца, которое может препятствовать передаче возбуждения с волоконца на синапс и далее на клетку, получило название пресинаптического торможения и считается одним из видов центрального торможения. При этом предполагается, что эта деполяризация создается в пресинаптических волоконцах путем специальной тормозящей импульсации по особым вставочным нейронам, синаптически оканчивающимся на пресинаптических волоконцах. Думают, что тормозящие нейроны выделением тормозящего медиатора производят деполяризацию пресинаптических волокон (Экклс и сотр., 64).
Мы не разделяем эту гипотезу прежде всего потому, что она требует наличия при каждом клеточном синапсе еще одного синапса на пресинаптическом волоконце, чтобы можно было вызвать полное торможение каждой
нервной клетки. В электронном микроскопе можно увидеть в некоторых случаях двойные синапсы, т. е. синапс на синапсе. Но никак нельзя утверждать, что дополнительный синапс производит торможение приклеточного синапса. Скорее всего надо допустить, что этот дополнительный синапс служит для возбуждения приклеточного. Когда в связи с постэмбриональным развитием или длительной периодической деятельностью нейрона количество синапсов сильно растет, они могут прилегать друг к другу и при этом дополнительный синапс будет действовать на приклеточный синапс возбуждающим образом. (Об умножении синапсов в связи с упражнением см. главу XVII об условных рефлексах.)
Выходящие из клетки дендритные токи производят деполяризацию мембраны сомы, а также начального сегмента аксона. Но эта деполяризация происходит в такой малой степени и с такой постепенностью ввиду своего медленного протекания, что обычно не вызывает распространяющегося возбуждения клетки. Так как через блокированные синапсы пирамидная клетка перестает получать постоянную импульсацию со стороны неспецифической облегчающей системы, а также со стороны возбужденных пирамидных и звездчатых нейронов, то существовавшая до этого фоновая деполяризация должна ослабнуть. А так как реполяризация клеточной мембраны значительно больше, чем та деполяризация, которая вызывается в ней дендритными токами, то клетка в общем претерпевает гиперполяризацию. Поэтому она становится менее возбудимой. Значит торможение клетки происходит в результате блокирования пресинаптических мембран.
Как указывалось выше, терминальные синапсы в первом слое в 3—4 раза крупнее, чем боковые синапсы, и содержат 3—4 митохондрии при отсутствии их в боковых синапсах. А так как при раздражении поверхности коры торможение ее происходит во время вызываемого им отрицательного медленного потенциала, то естественно заключить, что данное торможение в основном обязано своим происхождением этому потенциалу терминальных синапсов первого слоя.
Это хорошо выявляется, если сочетать электрическое раздражение коры в какой-либо первичной зоне с раздражением соответствующего рецептора, например, слуховую зону и слуховой рецептор. Как показано на рис. 37, во время раздражения слуховой зоны с усилением отрицательного потенциала происходит угнетение спонтанной электрической активности наряду с угнетением первичных ответов от слухового раздражения. Такое же угнетение наблюдается при раздражении коры, т. е. во время дендритных отрицательных потенциалов, в отношении отрицательных потенциалов нейронов /// слоя, вызванных через каллозальные волокна раздражением симметричной точки другого полушария (Чанг, 44).
Итак, при возбуждении аксодендритных синапсов в дендритах возникают локальные потенциалы. Вызванные ими токи без особого декремента проникают е тело клетки, а отсюда выходят через ближайшие синаптические участки, где электропроводность немного больше, чеммежду синапсами. Выйдяиз клетки, токи проникают в синаптические окончания и производят их анэлектротоническую блокаду, а выходя из синаптического окончания, вызывают деполяризацию пресинаптического волоконца. Вследствие всего этого приходящие по афферентам нервные импульсы не в состоянии подействовать на клетку. При этом клетка может гиперполяризоваться, ибо несмотря на некоторую деполяризацию постсинаптической мембраны в связи с выходом дендритных токов из клетки, благодаря блокированию синапсов в клетке исчезает та деполяризация, которая обычно вызывается под влиянием постоянной импульсации со стороны других клеток коры и подкорки.
Выше мы пришли к выводу, что в пирамидных клетках электрические потенциалы своими токами, распространяющимися по дендритам электротонически, производят анэлектротоническую блокаду дендритных синапсов и тем самым вызывают торможение дендритов. Сейчас мы сделали аналогичное заключение относительно торможения клеток в пирамидных нейронах.
Следовательно, дендриты и тело клетки в пирамидных нейронах находятся в реципрокных отношениях: усиленная активация клетки приводит к торможению дендритов, а усиленная активация дендритов, наоборот,— к торможению клетки.
По наблюдениям Полякова и других, известно, что коллатерали аксонов пирамидных нейронов главным образом пересекают дендриты соседних пирамидных нейронов или контактируют с ними. Это видно на рис. 38. С этим согласуется и физиологическое наблюдение. По данным Вулси и Чанга (45), при антидромном раздражении пирамидных путей в области продолговатого мозга, в коре мозга возникают медленные потенциалы, что указывает
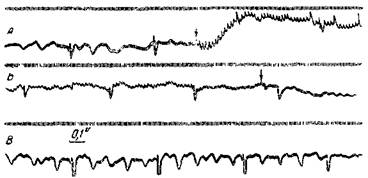
Рис. 37. Угнетение «спонтанной» электрической активности и «первичных ответов» от звуковых толчков при раздражении поверхности коры
Кошка под нембуталовым наркозом. «Активный» отводящий электрод расположен на среднейэктосильвиевой извилине; второй отводящий электрод на сигмоидальной извилине; раздражающие электроды — на поверхности средней эктосильвиевой извилины на расстоянии 4 мм от отводящего электрода. Усилитель с большой постоянной времени. А — эффекты звуковых толчков с частотой 5 в сек. и затем эффект присоединенного раздражения коры (частота — 50 в сек., интенсивность — 12 е); Б — продолжение комбинаций раздражений через 1,2 сек. и прекращение раздражения мозга при непрекращающемся звуковом раздражении; В— непосредственное продолжение Б (Беритов и Ройтбак)
на активацию дендритов коллатералями раздражаемых пирамидных нейронов. Мы находим, что когда возбуждается определенная группа пирамидных нейронов и одновременно через их коллатерали активируются дендриты ближайших пирамидных нейронов, то, согласно данной нашей концепции торможения, должно происходить торможение этих пирамидных нейронов (см. рис. 40). Этим путем осуществляется локализирование производимой корковой реакции.
Коллатерали аксона пирамидного нейрона могут оканчиваться и на собственных дендритах (см. рис. 38). Активация этих дендритов, как указывалось, должна вызывать торможение клетки. Но в определенных случаях, когда клетка только что возбудилась и деполяризация клеточной мембраны значительна, дендритный ток, возникший в базальных дендритах путем импульсации через коллатерали данного же нейрона, может так усилить деполяризацию близлежащего начального участка аксона, что вызывает распространяющееся возбуждение. При этом, конечно, дендритный ток, выходя из клетки также через синапсы, будет в некоторой мере блокировать пресинаптические мембраны. Однако это обстоятельство может не мешать означенному деполяризующему действию на начальный участок аксона. Возможно, что возбуждение с правильным и высоким ритмом 100—200 в сек. и выше, которое иногда наблюдал Джаспер при отведении от пирамидных клеток, в какой-то мере зависело от такого самовозбуждения клетки через свои коллатерали.
На рис. 39 дается схема распространения дендритных электрических токов в пирамидном нейроне при раздражении корковой поверхности и производимого ими торможения соседней пирамидной клетки.
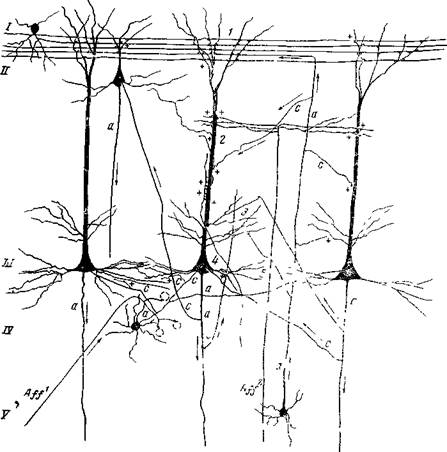
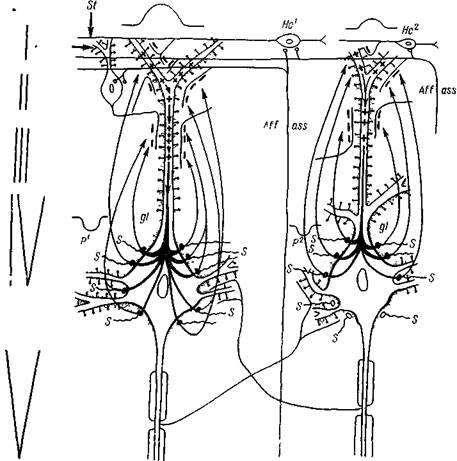
Рис. 38. Схема связей афферентных волокон со звездчатыми и пирамидными клеткам!Гкоры мозга
В I слое 'коры с верхушечными дендритами пирамидных нейронов ^контактируют ' горизонтальные аксоны клеток I слоя и аксон асссоциационной клетки Мартинотти. С верхушечными дендритами во II и III слоях контактируют афференты, приходящие из других отделов нервной системы, аксоны клеток Мартинотти и множество других аксонов. С ветвлениями дендрита в его нижней трети (3)| контактируют коллатерали аксонов клеток Мартинотти, коллатерали аксона исходной пирамидной клетки (контакт на себя). С телом (4) пирамидных клеток контактируют аксоны звездчатых клеток, коллатерали аксонов других пирамидных клеток. С базальвыми дендритами (5) контактируют аксоны звездчатых клеток* Каждая короткоаксонная звездчатая клетка контактирует с несколькими пирамидными клетками. А//' — афферент из подкорковых образований, приходящий в IV слой и контактирующий с коро ткоаксонными звездчатыми клетками; А//2 — афферент, приходящий в верхние отделы III слоя, контактирующий с дендритами пирамидных клеток; а — аксон; с — коллатераль аксона. Стрелками обозначено направ» ление передачи возбуждения.
Схему составила Школьник-Яррос (1958) на основании изучения препаратов своих собственных и Г. И. Полякова, импрегнированных хромосеребряным методом
Как видно из рисунка и как вытекает из всей нашей гипотезы, дендритное торможение клетки может происходить до некоторой степени благодаря блокаде части синапсов, наиболее близких к активному дендриту. Следовательно, такая пирамидная клетка может быть возбуждена через другие, более отдаленные синапсы.
На рис. 40 дается схема возбуждения пирамидной клетки и ее возбуждающего действия на базалыше дендриты соседнего пирамидного нейрона. Здесь же приводятся пути распространения электрических токов, производящих блокаду синапсов на дендритах данной клетки и на теле и дендритах соседней пирамидной клетки.
Дата добавления: 2017-01-13; просмотров: 929;