Локальное возбуждение дендритов.
Дендриты пирамидных нейронов приходят в активное состояние при возбуждении прилегающих к ним нервных окончаний. При этом в дендритах возникают длительные потенциалы около 10 мсек. Эти потенциалы локальны, не распространяются по дендриту и не доходят до клетки. Этого не бывает даже если в дендритах возникает суммированный потенциал вследствие возбуждения многих дендритных синапсов. Известно, что при локальном раздражении коры головного мозга, когда верхушечные дендриты пирамидных клеток возбуждаются и от поверхности коры отводятся значительные отрицательные потенциалы, сами пирамидные клетки не возбуждаются (Эдриан, 40). Можно предположить, что в основе этого явления лежат такие же особенности биохимических процессов, которые наблюдал Португалов (41) в дендритах двигательных нейронов спинного мозга. Именно активность сукциндегидразы в дендритах несколько слабее, чем в теле клетки и в зоне синапсов; кроме того, в дендритах по мере удаления от клетки содержание аминокислот — триптофана и гистамина уменьшается сравнительно с цитоплазмой тела клетки. Следовательно, можно предположить, что одной из причин сильного декремента в распространении локального процесса возбуждения в дендритах является малая интенсивность протекающих здесь ферментативных процессов.
При локальном электрическом раздражении коры возникающий здесь медленный отрицательный потенциал распространяется по поверхности коры на небольшом участке от места раздражения — не дальше 5—8 мм. Амплитуда потенциала падает и уже на расстоянии 10 мм равняется нулю. Но в отсутствие наркоза или во время очень легкого наркоза при локальном раздражении поверхностного слоя медленные потенциалы появляются по всей коре, но только они иного характера.
Как видно из рис. 32 поблизости от раздражаемого участка отводится главным образом отрицательный потенциал, а в более или менее удаленных участках, а также в противоположном полушарии отводятся положительные потенциалы. Отрицательный потенциал обычно появляется здесь после положительной фазы (Ройтбак, 34; Берне, 35; Чанг, 10).
Амплитуда отводимых отрицательных потенциалов градуально зависит от силы раздражения. С усилением раздражения (в 6 раз выше порогового) она быстро достигает максимума. Скрытый период отрицательных потенциалов, отводимых на разных расстояниях от раздражаемого участка, разный. На расстоянии 1,5—2 мм он наступает через 2—2,7 мсек. после момента раздражения. С удалением отводящего электрода амплитуда отрицательного
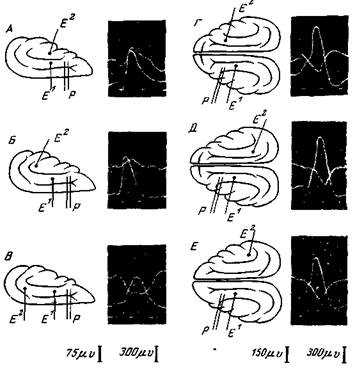
Рис. 32. Распространение активности из раздражаемого пункта по коре больших полушарий кошки.
Во всех опытах раздражается один и тот же пункт переднего полюса gyr suprasylvius; интенсивность раздражения 25 в, продолжительность раздражающего стимула 0,5 мсек. Первый отводящий электрод (Е1) расположен на gyp. suprasylvius на расстоянии 7 мм от раздражающих электродов (Р), на всех осциллограммах нижние кривые — потенциалы от Е1. Второй отводящий электрод (Е2) переставляется на разные участки коры А — на g. ectosylvius, передний ее полюс, В—на g. ectosyivius, задний ее полюс, В — на задний полюс g suprasylvius, Г— на g suprasylvius противоположного полушария в месте, симметричном месту раздражения, Д—на задний полюс g. suprasylvius противоположной стороны, Е — на задний полюс g. ectosylvius противоположной стороны (Ройтбак)
потенциала уменьшается, а скрытый период увеличивается (Берне, 35). Скорость распространения этого процесса очень мала — 0,5—0,8 м в сек. (Ройтбак, 34, 36).
Есть много оснований утверждать, что отрицательный потенциал, отводимый от поверхности коры около раздражаемого участка, возникает от развития в дендритах постсинаптических потенциалов в области крупных терминальных синапсов. Так как при раздражении поверхности коры отрицательные медленные потенциалы вызываются и при очень глубоком наркозе, когда возможность возбуждения нервных клеток исключена, да и клеточных элементов в первом слое очень мало, есть основания утверждать, что эти отрицательные потенциалы относятся к терминальным синаптическим участкам апикальных дендритов пирамидных нейронов. Причем, эти постсинаптические процессы вызываются не непосредственным раздражением дендритов, а через возбуждение нервных волокон первого слоя, имеющих с ними синаптические контакты. Тот факт, что вызванный одним электрическим стимулом данный отрицательный потенциал имеет продолжительность около 20 мсек, лучше всего свидетельствует, что он не может принадлежать ни нервным волокнам, продолжительность локального процесса в которых значительно меньше, ни глиальным элементам, продолжительность активирования которых, как известно, измеряется секундами (Ройгбак, 37).
В данном случае распространение отрицательного потенциала обусловливается распространением возбуждения по тем нервным волокнам, которые раздражаются электрическим током. В первом слое преобладают тонкие волокна вроде волокон группы С с малой скоростью проведения возбуждения. А так как эти волокна проходят по первому слою разные расстояния, то производимое ими активирование дендритов должно падать с расстоянием, т. е. соответственно уменьшению количества возбужденных волокон (Экклс, 38).
Но, возможно, что значительную роль здесь играет и то обстоятельство, что в особенно тонких нервных волокнах возбуждение распространяется с декрементом (Ройтбак, 36).
Локальное возбуждение в дендритах ни при каких силах раздражения не переходит в распространяющееся возбуждение. Это видно из того, что при углублении отводящего электрода под / и 77 слой, сколько-нибудь значительные отрицательные потенциалы перестают отводиться. Значит, возникшее в апикальных дендритах первого слоя возбуждение не распространяется по стволу дендрита и фактически ограничивается тем участком, где оно возникает. Кроме того, известно, что когда на поверхности коры возникает отрицательный потенциал, то на глубине 1,3 мм, где лежат клеточные тела и аксоны пирамидных клеток, быстрые колебания никогда не регистрируются (Берне, 35). Более того, когда дендриты ассоциационных нейронов возбуждаются при раздражении микроэлектродом, введенным в кору на глубину 0,3 мм (II слой), то разряда импульсов в их аксонах не происходит (Клер и Бишоп, 39). В связи с этим понятно, что сравнительно умеренное раздражение, приложенное к коре в двигательной зоне конечностей, приводит к появлению отрицательного потенциала, но не сопровождается сокращением мышц конечностей (Эдриан, 40).
В основе декремента возбуждения, как указывалось выше по Португалову (41), возможно, лежат особого рода биохимические процессы в двигательных нейронах спинного мозга: активность сукциндегидразы в дендритах двигательных клеток несколько слабее, чем в теле клетки и в зоне ее синапсов; кроме того, в дендритах, по мере удаления от тела клетки, содержание аминокислот — триптофана, тирозина и гистамина уменьшается по сравнению с цитоплазмой клетки и ограничивается в основном нейрофибриллами.
Итак, импульсы возбуждения, притекающие к верхушечным дендритам, вызывают в них настолько слабое местное возбуждение, которое к тому же настолько сильно декрементирует при своем распространении, что, можно сказать, оно ограничивается синаптическим участком. Поэтому оно не может достигнуть клетки и приводить к возникновению в ней распространяющегося возбуждения.
Но ток, вызванный этими потенциалами, электротонически достигает клетки и должен выходить из нее через синаптические участки и затем через аксонный холмик. Это приводит к некоторой деполяризации постсинаптических мембран и аксонного холмика, чем достигается облегчение последующего возбуждения клетки. Проходя через пресинаптические мебраны, дендритный электротонический ток будет производить их анэлектротоническую блокаду. Это, наоборот, затруднит передачу возбуждения к клетке и может вызвать смещение потенциала в сторону гиперполяризации вследствие прекращения всякого рода доступа межцентральных и афферентных импульсов к клетке.
Мы выше дали характеристику функциональной деятельности верхушечных дендритов пирамидных нейронов. Должно быть, она типична и для базальных дендритов, и в их аксодендритных синаптических участках возникающее возбуждение не должно достигать тела клетки. А возникающий при этом электротонический ток будет также действовать на клетку, как электротонический ток апикальных дендригов.
Дата добавления: 2017-01-13; просмотров: 1106;