Обгрунтуйте методи розвитку та контролю силових якостей спортсмена. 4 страница

Рис. 4. 3. Схематичне зображення м'яза:
М'яз (А) складається з м'язових волокон (Б), кожне з них ділиться на міофібрили (В). Міофібрила (Г) складається з товстих та тонких міофіламентів (Д). На рисунку показано один саркомер, обмежений з двох І боків лініями. І — ізотропний диск, А — анізотропний диск, Н — ділянка з меншою анізотропністю. Поперечний розріз міофібрили (Е) дає уяву про гексагональне розташування товстих та тонких міофіламентів.
Просторове розташування протофібрил надає скелетним м'язам характерного вигляду смугастості, яку добре видно під світловим мікроскопом.
Тонкі мембрани (2), до яких прикріплюються тонкі протофібрили, поділяють міофібрили на структурні одиниці — саркомери. В саркомерах нитки актину знаходяться між нитками міозину. В ділянці перекриття їх вони з'єднуються поперечними містками. Поперечна смугастість є результатом чергування ділянок саркомера, з сильним і слабким заломленням променів. У тій частині" саркомера, де знаходяться тільки тонкі протофібрили (диск /), заломлення променів невелике. Під мікроскопом ці ділянки саркомера здаються світлими. В місцях перекриття товстих і тонких протофібрил (диск А) заломлення променів подвійне, вони здаються темними. Всередині диска А тонких протофібрил немає, тому там розташована світла зона — Н. Міофібрили групуються в колонки по 4 ... 20 в кожній. Колонки оточені системою трубочок і міхурців, яку називають саркоплазматичною сіткою (рис. 4). Трубочки, які тягнуться вздовж міофібрил, утворюють так звану Т -систему. Біля 2-мембран трубочки Т-системи розширюються, утворюючи латеральні цистерни, в яких містяться іони кальцію. Поздовжні трубочки з'єднуються між собою за допомогою поперечних трубочок І-системи. Системи поздовжніх (Т-система) і поперечних (І-система) канальців забезпечують передачу збудження до м'язових волокон і перехід від збудження до скорочення.

Рис. 4. 4. Схематичне зображення структури саркоплазматичного ретикулума двох фібрил з м'яза
В обох фібрилах повністю зображено лише один саркомер; зліва — саркомер з інтактним ретикулумом, справа ретикулум частково відділено, для того, щоб показати міофібрили. Можна бачити, що поперечні елементи ретикулума беруть початок у плазмалемі (//). Темні потовщення на кінцях поздовжніх пухирців — це кінцеві цистерни тріад, розташовані в цьому випадку на границі А- та І-дисків, С — саркоплазма, М — мітохондрії, О — основна речовина.
У м'язах ссавців виділяють три типи м'язових волокон — А, В і С. Волокна типу А — світлі, з малим вмістом міоглобіну, високою здатністю використовувати глікоген як джерело енергії, низьким рівнем ліпідного обміну і процесів окислення. Волокна типу С, навпаки, темні, багаті на мітохондрії, з низькою здатністю розщеплювати глікоген, високим рівнем процесів окислення і великим вмістом міоглобіну. Волокна типу В за рядом властивостей займають проміжне положення. Спектр волокна від світлих до темних відповідає поступовому переходу здатності їх від анаеробного (безкисневого) розщеплення вуглеводів до аеробного обміну. Функціональне значення диференціації м'язових волокон полягає в їхньому пристосуванні до слабких і тривалих (фазичних) напружень. Білі м'язові волокна мають повільний, тонічний, а темні м'язові волокна швидкий, фазичний характер скорочення [11, 16].
Біомеханічні властивості м'язів
До біомеханічних властивостей м'язів відносять скоротність, пружність, жорсткість, міцність і релаксацію.
Скоротність– це здатність м'яза скорочуватися при збудженні. В результаті скорочення відбувається укорочення м'яза і виникає сила тяги.
Пружністьм'яза полягає в його здатності відновлювати первинну довжину після усунення деформуючої сили. Існування пружних властивостей пояснюється тим, що при розтягуванні в м'язі виникає енергія пружної деформації. При цьому м'яз можна порівняти з пружиною: чим сильніше розтягнута пружина, тим більша енергія в ній запасена.
Жорсткість– це здатність протидіяти силам, що прикладаються. Коефіцієнт жорсткості визначається як відношення приросту поновлюючої сили до приросту довжини м'яза під дією зовнішньої сили:
Кж=DF/Dl (Н/м)
Величина, зворотна жорсткості, називається податливістюм'яза. Коефіцієнт податливості:
Кп=Dl /DF (м/Н)
– показує, наскільки подовжиться м'яз при зміні зовнішньої сили. Наприклад, податливість згинача передпліччя близька до 1 мм/Н.
Міцністьм'яза оцінюється величиною розтягуючої сили, при якій відбувається розрив м'яза. Сила, при якій відбувається розрив м'яза складає від 0.1 до 0.3 Н/мм2. Межа міцності сухожилків на два порядки величини більше і складає 50 Н/мм2. Проте, при дуже швидких рухах можливий розрив більш міцного сухожилка, а м'яз залишається цілим, встигнувши амортизувати.
Релаксація– властивість м'яза, що виявляється в поступовому зменшенні сили тяги при постійній довжині м'яза. Релаксація виявляється, наприклад, при стрибку вгору, якщо під час глибокого присідання спортсмен робить паузу. Чим пауза триваліше, тим сила відштовхування і висота виплигування менше.
4.4. Фізіологічна характеристика рухових одиниць м'язів
Головним структурно-функціональним елементом нервово-м'язового апарата є рухова одиниця (РО), її утворюють: 1) тіло мотонейрона, яке знаходиться в передньому розі спинного мозку, або в руховому ядрі головного мозку; 2) аксон цього нейрона (один довгий відросток); 3) кінцеві розгалуження аксона; 4) нервово-м'язові синапси; 5) м'язові волокна, з якими з'єднується аксон. Усі РО організму складають нервово-м'язовий апарат. М'язове волокно є структурною одиницею м'яза, а РО - структурно-функціональною одиницею нервово-м'язового апарата. Нервово-м'язовий апарат не слід ототожнювати з опорно-руховим апаратом людини, до складу якого окрім рухових одиниць, входять кістки скелету, суглоби і зв'язки.
Рухові одиниці відрізняються між собою розмірами тіл мотонейронів, товщиною аксонів, а також числом і типом м'язових волокон. У зв'язку з цим виділяють великі і малі РО. Кількість м'язових волокон у них коливається від декількох десятків одиниць (м'язи ока) до декількох тисяч (литковий м'яз). Малі РО переважно входять до складу м'язів, які забезпечують швидкі і точні рухи (м'язи очного яблука, пальців кисті, дрібні м'язи обличчя ). В інших м'язах малі РО забезпечують повільні рухи. Великі РО, як правило, входять до складу м'язів тулуба і кінцівок, які здійснюють як швидкі, так і повільні рухи. Великі РО є переважно високопороговими Їх активація можлива лише при виконанні фізичних вправ із значним напруженням м'язів. В цілому ж в кожному м'язі є різні за розміром РО, що зумовлює можливість розвитку ними різної величини напруження.
Функціонально РО поділяють на повільні (тип І) і швидкі -(тип II). Всі м'язові волокна, які входять до складу діючої РО, за своїми властивостями ідентичні. Кількісне співвідношення м'язових волокон цих двох типів РО має генетичне походження і змінюється в онтогенезі за спадковою програмою. У дітей дошкільного і молодшого шкільного віку основна маса скелетних м'язів складається з волокон типу І. В період статевого дозрівання під впливом статевих гормонів, особливо тестостерону, в скелетних м'язах настають значні зміни в розвитку м'язових волокон. Починають швидко збільшуватись у поперечному розрізі і в кількості волокна типу II. Абсолютна кількість волокон типу і при цьому залишається незмінною (Д.А.Фарбер та ін., 1990).
Таблиця.4.1.
Структурна і функціональна характеристика різних типів м 'язових волокон
| Характеристика | Тип волокна | ||
| І | ІІ А | ІІ Б | |
| Кількість волокон на матонейтрон | 10-180 | 300-800 | 300-800 |
| Розмір мотонейрона | невеликий | великий | великий |
| Швидкість нервової провідності | невелика | велика | велика |
| Швидкість скорочення, мс | |||
| Тип міозинової -АТФ-фази | повільний | швидкий | швидкий |
| Сила рухової одиниці | невелика | велика | велика |
| Аеробна здатність | висока | середня | низька |
| Окислювально-гліколітична здатність | низька | висока | висока |
Сила, швидкість скорочень і витривалість повільних і швидких м'язових волокон різні, що зумовлено їх морфологічними і біохімічними особливостями. Повільні м'язові волокна («червоні» м'язи) мають багато капілярів, міоглобіну і мітохондрій. Вони повільно стомлюються, забезпечуючи довготривалі м'язові скорочення порівняно невеликої сили (стаєрський і марафонський біг, лижні гонки, велосипедний спорт тощо). Чим більше генетично обумовлених повільних волокон, тим вищий спадковий показник максимального споживання кисню. Робоча гіпертрофія повільних волокон лежить в основі розвитку витривалості.
Швидкі м'язові волокна (тип II) називаються гліколітичними, це — «білі» м'язи. На відміну від повільних, окислювальних волокон, вони працюють в безкисневому (анаеробному) режимі. Розвиваючи велику силу статичних і динамічних скорочень швидкі м'язові волокна зумовлюють «вибухову» силу і високу швидкість рухів (спринтерський біг, стрибки, підняття штанги).
У процесі спортивних тренувань на витривалість можна досягнути перетворення частини швидких чисто гліколітичних анаеробних білих волокон (тип II) у другу частину швидких волокон - у волокна окислювально-гліколічні цього ж типу (ІІА), які можуть скорочуватись сильно, інтенсивно, протягом тривалого часу в аеробних і анаеробних умовах, повільно стомлюючись (табл.4.2). Такі перетворення можливі тільки в межах II типу м'язових волокон. Таким чином, поруч з поділом волокна на І і II типи вчені стали розрізняти м'язові волокна двох видів типу II: підтип - II А і підтип II Б. Волокна підтипу IIА за своїми властивостями наближаються до повільних волокон типу І.Вони окисно-гліколітичні, тобто мають можливість працювати як волокна типу І (повільні), і як волокна типу II Б (швидкі). Волокна підтипу II А здатні функціонувати не тільки в анаеробних, а й в аеробних умовах, пристосовуючись до виконання довготривалих фізичних вправ великої сили і великої швидкості.
Дослідженнями вчених встановлено, що спринтери відрізняються від стайєрів кількістю швидких (білих) м'язових волокон. У спринтерів - 80-85% швидких і 20-15% повільних (червоних) волокон, а у стайєрів 80-90% повільних і 10-15% швидких волокон. Отже, на короткі дистанції (швидкісно - силова робота) доцільно спеціалізувати осіб, у м'язах яких міститься не більше 30% м'язових волокон типу І, а на довгі дистанції (робота на витривалість) - спортсменів, м'язи яких містять 60% м'язових волокон цього типу. Доведено, що спортсмени, в м'язах яких переважають повільні м'язові волокна, не можуть досягнути значних результатів у спринті, тому що кількісне співвідношення волокон типу 1 і волокон типу II не змінюється в процесі фізичних тренувань. Цей факт необхідно враховувати в практиці фізичного виховання, зокрема при вирішенні питань спортивної орієнтації школярів.
Електроміограма (ЕМГ). Скорочення м'язового волокна супроводжується появою потенціалу дії. При активації багатьох рухових одиниць з поверхні м'яза реєструють сумарну біоелектричну активність рухових одиниць. Сум'арна електрична активність рефлекторного скорочення при слабкому напруженні складається із нерегулярних коливань потенціалу малої амплітуди. Із збільшенням сили скорочення коливання стають ритмічними, амплітуда їх збільшується. Основна частота електроміограми коливається в межах 20 ... 50 імпульсів за 1 секунду, на яку накладаються нерегулярні хвилі. Між сумарною амплітудою електроміограми (інтегрована ЕМГ) і силою скорочення м'яза в межах від слабких до субмаксимальних напружень існує лінійна залежність [11, 16].
4.5. Форми і типи м'язових скорочень
В залежності від характеру зміни довжини м'яза, що скорочується, розрізняють динамічну, статичну і ауксотонічну форми м'язового скорочення.
Динамічнимназивається скорочення м'яза, яке викликає зміну його довжини. Таке скорочення лежить в основі динамічної роботи. Динамічні скорочення бувають двох типів: концентричні і ексцентричні.
Коли зовнішнє навантаження менше, ніж напруження м'яза, що скорочується, м'яз, скорочуючись, вкорочується, викликаючи рух, це концентричний тип скорочення. Якщо ж скорочення м'яза здійснюється при сталому напруженні, то його називають ізотонічним (ізос - рівний, тонус - напруження). Якщо зовнішнє навантаження більше, ніж напруження м'яза під час скорочення говорять про ексцентричний або міо-метричний тип скорочення. Такий м'яз розтягується в час скорочення. В природних умовах даний тип скорочення може спостерігатися при роботі м'язів в уступаючому режимі (опускання вантажу, робота м'язів-розгиначів у час скорочення м'язів-згиначів). Динамічні вправи, які лежать в основі динамічної роботи, мають фізичне вираження (кгм, Вт, Дж, ккал), для них може бути визначений коефіцієнт корисної дії.
Скорочення м'язів, при яких вони розвивають напруження, але не змінюють своєї довжини, називають статичними.
Такі скорочення забезпечують підтримання тіла в просторі, вони направлені на протидію земному тяжінню і сприяють збереженню відповідної пози тіла. Підтримання природної пози спортсмена здійснюється енергоекономічними, мало-втомлюваними тонічними напруженнями м'язів. Більшість же статичних вправ (положень, поз), які зустрічаються у спортивній практиці, пов'язані з тетанічними напруженнями м'язів
Скорочення, при яких змінюється довжина і напруження м'язів, називають ауксотонічним.
Така форма скорочень м'язів зустрічається найчастіше, адже в реальних умовах діяльності людини чисто ізометричні і чисто ізотонічні скорочення практично не зустрічаються. Узагальнена характеристика форм і типів м'язових скорочень подана в табл. 4.2.
У відповідності з типом скорочення м'язів, які забезпечують виконання даної вправи, усі фізичні вправи поділяються на статичні та динамічні. Відповідно і будь-яка фізична робота в крайніх значеннях може бути динамічною і статичною. Частіше статична робота є тимчасовим елементом в одному з циклів динамічної роботи.
Таблиця 4.2..
Характеристика форм і типів м 'язових скорочень
| Форми скорочення м'язів | Тип скорочення | Зовнішнє навантаження | Зовнішня робота |
| 1 .Динамічна | концентричний (ізотонічний) | менше, ніж напруження м'язів | позитивна |
| ексцентричний | більше, ніж напруження м'язів | негативна | |
| 2. Статична | ізометричний | дорівнює напруженню м'язів | рівна нулю |
При динамічному скороченні виконується зовнішня робота: при концентричному скороченні - позитивна, при ексцентричному - негативна. Співвідношення вираженості динамічних і статичних скорочень у виконанні даної вправи дозволяє умовно поділити їх на долаючі (підняття вантажу), підтримуючі (утримання вантажу) та уступаючі (опускання вантажу). Долаючі вправи характеризуються концентричним (ізотонічним) скороченням м'язів внаслідок їх вкорочення. Уступаючі скорочення - ексцентричні. В них зовнішня сила, яка діє на м'язи, більша за силу, яку розвиває працюючий м'яз. З долаючими та уступаючими вправами пов'язана уява про позитивну та негативну роботу.
4.6. Режими скорочення м'язових волокон
В залежності від частоти імпульсації мотонейрона м'язові волокна рухової одиниці (РО) можуть працювати в двох режимах - в режимі поодинокого скорочення і в режимі тетанічного скорочення.
Режим поодиноких скорочень виникає тоді, коли інтервал між суміжними імпульсами мотонейрона дорівнює, або дещо більшій тривалості поодинокого скорочення іннервованих ним м'язових волокон.
В кривій поодинокого ізометричного скорочення м'яза виділяють три фази: латентний (прихований) період збудження, фаза підйому напруження і фаза розслаблення. При ізотонічному скороченні ці фази позначаються дещо інакше: латентний період збудження, фаза вкорочення, фаза видовження.
Тонічне напруження м'язів. Здатність скелетної мускулатури тривало, стійко підтримувати скорочення (напруження) називається тонусом, або тонічним напруженням.
Тонус м'язів має рефлекторне походження. Його виникнення і підтримання зумовлене наявністю в м'язах тонічно напружених рухових одиниць, які постійно контролюються нервовими центрами. В свою чергу, тонус нервових центрів підтримується по зворотних зв'язках аферентними імпульсами, що йдуть від пропріорецепторів. М'язовий тонус є основою підтримання необхідної пози тіла в умовах постійної дії сил земного тяжіння.
При руховій діяльності звичайно до м'язових волокон надходить серія наступних нервових імпульсів, які викликають тривале тетанічне скорочення м'язів - тетанус. Тетанічні скорочення властиві тільки скелетним м'язам і не характерні для гладеньких м'язів внутрішніх органів та поперечносмугастих м'язів серця, що пояснюється тривалим рефрактивним періодом. Тетанічні скорочення, в порівнянні з поодинокими, більш тривалі та сильні.
Тетанус викликається сумацією поодиноких м'язових скорочень, коли повторний нервовий імпульс надходить до м'яза раніше, ніж повністю завершиться його поодиноке скорочення. За таких умов відбувається накладання однієї хвилі поодиноких скорочень на другу (явище суперпозиції). Якщо ж кожний наступний імпульс надходить до м'яза в той момент, коли він починає розслаблюватись, то виникає зубчастий неповний тетанус, якщо ж повторний імпульс приходить в період його скорочення - виникає суцільний (повний) або гладенький тетанус. Суцільний тетанус дає більш сильне скорочення, ніж зубчастий тетанус. Частота імпульсів, яка необхідна для виникнення тетанусу, обернено пропорційна швидкості поодиноких скорочень. Для виникнення суцільного тетанусу в повільних РО достатня частота імпульсів 25 за 1 с, а для швидких РО-50 імпульсів за 1 с і більше (Рис 4.5.)
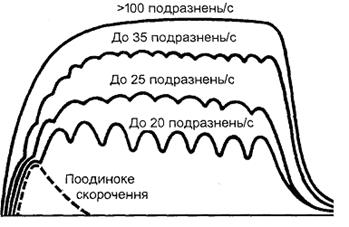
Рис. 4.5. Формування тетануса в залежності від частоти подразнення
Контрактура. Поряд з тетанічним скорочення м'язів зустрічається ще одна різновидність тривалого скорочення м'язів - контрактура.
Контрактура - це локальне і сильне довготривале скорочення м'язових волокон. Воно характеризується стійким напруженням (стягуванням) м'яза з сильно сповільненим розслабленням.
Контрактура буває природженою (природжені різкі обмеження рухомості внаслідок недорозвинутості м'язів і суглобів) і набутою. Набуті контрактури виникають при порушеннях функцій нервової системи, а також в умовах дії надмірного за силою больового, або температурного подразника. Професійні контрактури зумовлені тривалою активністю тих груп м'язів, які інтенсивно використовуються в даній професії. Контрактура при стомленні характеризується збільшенням тривалості періоду скорочення та розслаблення м'язів. У даному випадку вона зумовлена змінами обміну речовин у м'язах.
4.7. Характеристичні криві м'язів
Крива довжини — напруження. М'язи мають певні механічні властивості, які впливають на скороченнях їх. Схематично м'яз можна уявити у вигляді системи, яка складається із скоротливих і еластичних пружних елементів (рис. 4.6.). Розрізняють два пружних компоненти. Один з них розташований послідовно із скоротливим елементом, другий — паралельно. Послідовний пружний компонент утворений сухожилками і поперечними місточками між нитками актину і міозину. Він відіграє певну роль у забезпеченні плавності рухів. Паралельний пружний компонент складається із прошарку сполучної тканини, що оточує м'яз, і знаходиться всередині його. При розтягненні еластичних елементів м'яза зростає його напруження.

Рис. 4.6. Схеми, що пояснюють механіку ізометричного та ізотонічного скорочення
Зверху: А — механічна модель м'язового волокна, що включає скоротливий компонент (СК) та пасивні пружні компоненти — послідовний пружний компонент (ПОПК) та паралельний пружний компонент (ППК); Б — схематичне зображення моделі ізометричного скорочення (F — загальна довжина м'яза, F’ — напруження спокою, F’’ — напруження при скороченні); В — графік зміни напруження під час ізометричного скорочення. Знизу: А — схематичне зображення моделі ізотонічного скорочення м'яза; Б — графік зміни напруги; В — довжина м'яза при ізотонічному скороченні. Видно, що спочатку при ізотонічному скороченні збільшується «внутрішня напруга», коли воно досягає максимального значення, починається вкорочення м'яза. Під час вкорочення напруга (F) залишається незмінною і дорівнює зовнішньому навантаженню (Р).
Напруження, яке розвиває м'яз при подразненні, залежить від його довжини. Аналізуючи графік кривої «довжина — напруження», можна виділити на ньому п'ять критичних точок. При надмірному розтягненні волокна довжина саркомера збільшується до 3,6 мкм, актинові і міозинові нитки втрачають поперечні зв'язки, і тому напруження не розвивається, воно дорівнює нулю. У міру скорочення саркомера нитки актину зчеплюються з міозином — напруження збільшується і при довжині саркомера 2,25 мкм, що дорівнює 1,2 його довжини в розслабленому стані, площа зщеплення між міозином і актином найбільша. Тому і напруження волокна при цій довжині саркомера найбільше. При вкороченні саркомера до 20 мкм (розслаблений стан волокна) величина напруження суттєво не змінюється. Далі вкорочення саркомера зменшує площу ниток актину і міозину, а, як наслідок, і ступінь напруження волокна. При довжині саркомера 1,25 мкм міозинові нитки впираються в мебрану і згинаються, волокно втрачає здатність розвивати напруження.
Крива «сили — швидкості». Швидкість скорочення є функцією навантаження: чим більше навантаження, тим менша швидкість скорочення. У граничних випадках при відсутності навантаження швидкість скорочення максимальна, а при максимальному ізометричному напруженні вона дорівнює нулю. Графічно крива «сили — швидкості» має вигляд експоненти (рис. 8) і може бути описана рівнянням А. Хілла:
U = b (Р0—Р) • (Р+а),
де U — швидкість скорочення,
Р — сила (навантаження),
Р0 — максимальне ізометричне напруження, яке може розвинути м'яз,
b — константа швидкості,
а — константа сили.
Константи а і b визначаються на основі експериментальних даних. Експоненціальна залежність між силою напруження м'яза і швидкістю його скорочення зумовлена неодночасною активацією швидких і повільних рухових одиниць. При невеликій силі напруження активуються швидкі рухові одиниці з великою швидкістю скорочення. При збільшенні навантаження на м'яз необхідна величина сили напруження досягається за рахунок включення повільних рухових одиниць. Це зумовлює різке зменшення швидкості скорочення м'яза [15, 4].
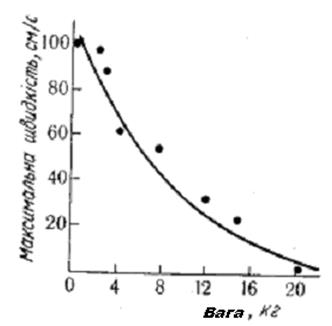
Рис. 4.7. Крива „сили - швидкості” мязового скорочення
4.8. Сила і робота м'язів
При збудженні м'язові волокна скорочуються і в них розвивається напруження. Величина максимального напруження під час збудження характеризує силу м'яза. Сила м'яза залежить від ряду факторів:
1) сили скорочення окремих його волокон;
2) кількості волокон у м'язі;
3) початкової довжини м'яза;
4) характеру нервових впливів на м'яз.
Результуюча силова дія м'язів на скелет зумовлюється процесами виникнення і регулювання сил тяги в самому м'язі, а також особливостями розташування м'язів на опорному апараті, геометричним прикріпленням їхніх сухожилків до кісток.
Встановлено, що одне м'язове волокно може розвинути напруження в 100... 200 мг. Товсті волокна розвивають більше напруження, ніж тонкі. Для порівняння сили м'язів визначають абсолютну їх силу. Абсолютна сила вимірюється в ньютонах на 1 см2 фізіологічного поперечника м'яза. Фізіологічний поперечник — це площа поперечного розрізу всіх волокон м'яза, проведеного перпендикулярно їх розташуванню. У перистих м'язах фізіологічний поперечник значно більший від анатомічного (площі м'яза за його найбільшим діаметром), тому вони сильніші, ніж м'язи з паралельним розміщенням волокон. Абсолютна сила м'язів людини характеризується такими величинами (в кілограмах на 1 см2): литковий м'яз — 5,9; згиначі плеча — 8,63; жувальні мязи—10,0; двоголовий м'яз плеча — 11,4; триголовий м'яз плеча — 16,8.
Регуляція напруження м'язів здійснюється в основному двома факторами: зміною числа активованих рухових одиниць і частотою нервових імпульсів. Чим більша кількість рухових одиниць збудження, тим більше напруження розвиває м'яз. Підвищення частоти імпульсів від мотонейронів також збільшує силу скорочення м'язів.
Робота м'язів. Під час скорочення м'язи виконують певну роботу. Розрізняють внутрішню роботу, пов'язану з витратами енергії на процеси, які забезпечують саме скорочення м'яза (розтягування еластичних елементів, проведення збудження, ре-синтез АТФ), і зовнішню роботу, під час якої енергія скорочення м'яза перетворюється на потенціальну або кінетичну енергію. Зовнішню роботу називають ще механічною роботою. Величина її (А) в найбільш простих випадках визначається добутком величини сили (F) на відстань її дії (s), тобто
А = FS
Наприклад, якщо важкоатлет прикладає силу у 150 Н для підняття штанги на відстань у 2 м, то його м'язи виконують роботу, що дорівнює 2940 Нм1.
Але дле це наближена цифра, оскільки не враховується робота, яка виконується при розгинанні тулуба, піднятті рук, підтриманні певної пози тіла. Визначити цю додаткову роботу досить важко. Величина механічної роботи, що виконується м'язом, залежить від навантаження м'яза. Ця залежність може бути продемонстрована графіком, зображеним на рис. 4.8., з якого видно, що максимальна робота досягається при середніх величинах навантаження [4, 5].
 .
.
Рис. 4.8. Залежність механічної ефективності мязової роботи від сили скорочення при ізометричному режимі
Встановлено, що величина зовнішньої роботи залежить від швидкості скорочення м'язів і максимальною вона є при середній швидкості скорочення. Ці залежності між величиною роботи м'яза і навантаженням та швидкістю його скорочення у відповідності носять назву закону середніх навантажень і швидкостей.
Більш точним і важливим показником ефективності роботи м'язів є коефіцієнт корисної дії (ККД), який є відношенням виконаної механічної роботи (А) до загальних енергетичних витрат (Q). Загальні енергетичні витрати складаються із витрат на механічну роботу і на утворення тепла (H), тобто Q=А + Н, звідки
ККД = А / Q
Як показують дослідження, ККД м'язів людини може досягати 25 ... 30%, тобто тільки 30% всієї енергії скорочення витрачається на механічну роботу, а 70% розсіюється у вигляді тепла. ККД також залежить від величини навантаження і швидкості скорочення. ККД максимальний при навантаженнях, що дорівнюють половині від максимальної сили м'язів і при швидкості скорочення 0,35 від максимальної.
Закони середніх навантажень і середніх швидкостей визначають продуктивність роботи рухового апарату і мають важливе значення для трудової і спортивної практики. У процесі тренування і росту дітей збільшуються функціональні можливості рухового апарату, збільшуються і абсолютні значення середніх навантажень і швидкостей. Тому величину середніх навантажень і швидкостей визначають експериментально в кожному окремому випадку (для дітей різного віку, для різних м'язів і різних рухів).
Дата добавления: 2016-06-13; просмотров: 2832;