Типы бластул и способы гаструляции. У оболочников, бесчерепных и низших позвоночных (анамний) образовавшаяся в результате дробления бластодерма разделяется в процессе гаструляции на два слоя —
См. пред. +
У оболочников, бесчерепных и низших позвоночных (анамний) образовавшаяся в результате дробления бластодерма разделяется в процессе гаструляции на два слоя — эпибласт ( ки-нобласт И. И. Мечникова, или верхний листок А. О. Ковалевского) и гипобласт ( фагоцитобласт Мечникова, или нижний листок Ковалевского). Первый дает начало эктодерме и отчасти мезодерме, а последний — энтодерме, мезодерме и хорде.
У высших позвоночных (амниот) от бластодермы в первой фазе гаструляции отщепляется первичная энтодерма, после чего бластодерма вступает во вторую фазу гаструляции, когда обособляются эпибласт, дающий, как и у анамний, эктодерму и отчасти мезодерму, и гипобласт, на этот раз образующий только хорду и мезодерму, так как энтодерма уже обособилась во время первой фазы.
НАРИСОВАТЬ КАРТУ ПРЕЗУМПТИВНЫХ ЗАЧ ПТ И АМФ
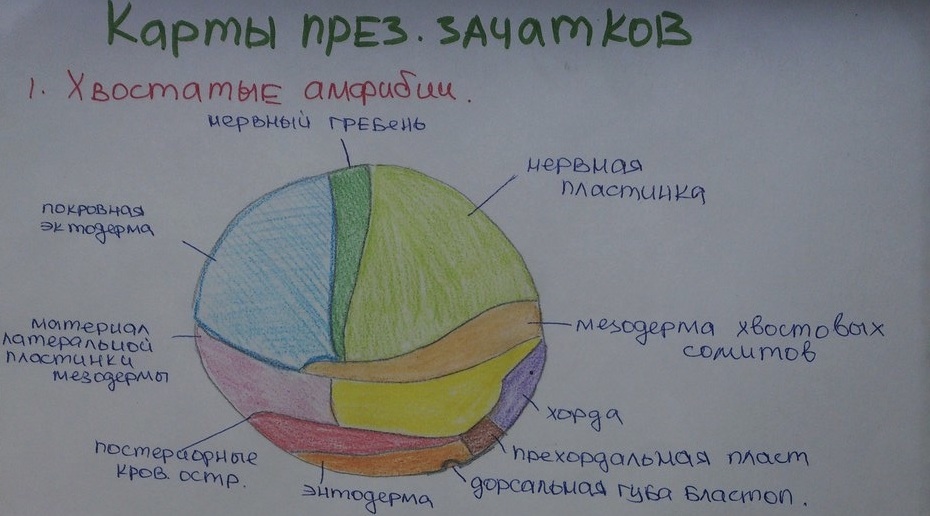
Амфибии:
Непрерывно углубляющаяся полость ямки инвагинации соответствует полости первичного кишечника (архентерона) ланцетника. Ее углубление обусловливается весьма характерным перемещением презумптивного материала в дорсальной (передней) и в латеральных (боковых) губах бластопора. Клетки в этих отделах начинают быстро размножаться и так же, как у ланцетника, иммигрируют внутрь ямки (архентерона), то есть, как принято говорить, происходит нарастание губ бластопора. Различие, но одновременно и общность передвижений этого материала с гаструляцией ланцетника показаны на приведенных рисунках. Точно так же, как и у ланцетника, хордомезодермальный материал нарастает на передней губе и через край бластопора перемещается внутрь, располагаясь по средней линии спины под эктодермальной полоской будущей медуллярной пластинки. Ход хордальной закладки и медуллярной пластинки определяет, таким образом, продольную ось удлиняющегося зародыша. Плечи ямки инвагинации, которая сначала имеет подковообразный вид, представляют собой латеральные губы бластопора; через них происходит нарастание презумптивного материала для мезодермы, который проникает под поверхностную эктодерму и перемещается по направлению к черепному концу эмбриона. При этом он располагается по бокам хордальной закладки, а краниально соединяется с головной мезодермой, берущей начало из области передней губы.
Таким образом как у ланцетника, так и у амфибий, образуется спинная сторона тела зародыша. (Сравни с предыдущей главой о гаструляции и нотогенезе ланцетника). Наконец, мезодермальный материал нарастает и перемещается и через край задней губы, тем самым ограничивая ее и замыкая до сих пор еще открытую заднюю границу бластопора. Эта мезодерма перемещается в хвостовом направлении, проникая под энтодерму и, в конце концов, соединяясь с остальной (парахордальной) мезодермой в единый средний зародышевый листок. Вместе с ней через отверстие бластопора медленно перемещается и громоздкая масса вегетативных клеток, которая вначале выступала из округлого бластопора в виде так называемой желточной пробки Рускони. Наряду с этой несовершенной инвагинацией, в процессе гаструляции наблюдается еще и новое явление, также обусловленное повышенным количеством желточного вещества в вегетативных макромерах. Пигментированная область бластодермы, соответствующая клеткам анимального полюса, из которых позднее развивается эктодерма, начинает в результате активного роста перемещаться по направлению к краям бластопора, то есть по направлению к вегетативному полюсу; таким образом, значительная часть будущих энтодермальных клеток, содержащих большое количество желтка, попадает во внутрь зародыша благодаря тому, что происходит их обрастание эктодермой. Этот сопутствующий процесс развития называется эпиболией; на более поздних стадиях филогенетического развития эпиболическое обрастание поверхностной эктодермы начинает превалировать над процессом инвагинации. При этих процессах в результате образования и расширения полости первичного кишечника масса вегетативных клеток, создающая закладку будущей энтодермы, поворачивается на 180 градусов и образует дно и боковые стенки архентерона. Подобным образом, как и у ланцетника, у земноводных позднее из этой энтодермы формируется примитивный, первичный, кишечник. Дорсальная крыша кишечной закладки сначала не замкнута, и в этом месте в его энтодерму проникает материал хорды и парахордальной мезодермы. В соответствии с развитием полости первичного кишечника, с углублением ямки инвагинации и с обрастанием губ бластопора презумптивным материалом во внутрь постепенно исчезает и полость бластоцеля.
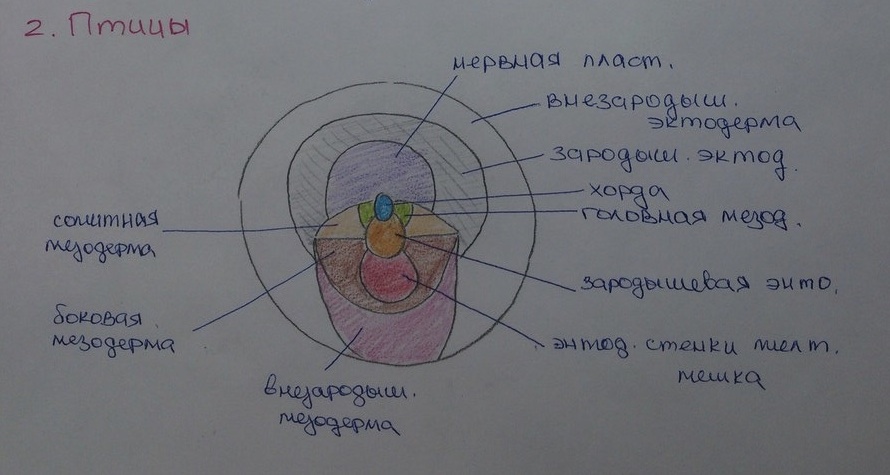
Птицы:
При дальнейшем развитии дискобластулы пресмыкающихся и птиц сначала возникает энтодерма, которая, однако, в данном случае уже не может развиваться посредством инвагинации и поэтому образуется иным, новым путем. В то время как у некоторых видов пресмыкающихся (например у черепах) и у некоторых рыб еще наблюдаются кое-какие черты инвагинирующего процесса, то у птиц инвагинация отсутствует полностью. Измененные условия существования, наряду с большим содержанием желтка, сделали у этих животных невозможным образование энтодермы путем инвагинации и обусловили возникновение принципиально иного вида гаструляции.
Энтодерма у пресмыкающихся и у птиц возникает в результат процесса, при котором происходит отделение клеток и клеточных совокупностей от бластодермы (отчасти и от каудального конца зародышевого щитка из отдела зародышевого валика) и их распределение в сплошной клеточный слой, благодаря чему первоначальная бластодерма расщепляется на поверхностную эктодерму и расположенную под ней энтодерму. Таким образом, гаструляция в данном случае происходит по новому способу — путем деляминации (расщепления) энтодермального слоя. У пресмыкающихся таким образом возникает двуслойный зародыш — дискогаструла, у птиц — эпигаструла. После окончания данного процесса развития наступает нотогенез, происходящий в области блестящей зоны (area pellucida), а именно в ее центральной части (в зародышевом щитке, то есть в эмбриональной зоне). У пресмыкающихся в задней части этого зародышевого щитка образуется узелковое утолщение эктодермы — примитивная или первичная ротовая пластинка. В этой пластинке клеточный материал начинает проникать вглубь, благодаря чему возникает карманоподобное образование — примитивный, первичный карман, проникающий между эктодермой и энтодермой. Полость этого кармана отчасти соответствует полости первичного кишечника, и ее можно считать несовершенным результатом процесса инвагинации. Первичный или иначе хордомезодермальный карман у пресмыкающихся растет в краниальном направлении, причем края отверстия в его полость соответствуют передней губе бластопора. Материал этой губы в процессе роста и подсовывания кармана попадает внутрь и дает начало хорде и головной мезодерме. Основание кармана, прилежащее к энтодерме, разрывается, и возникает свободное сообщение полости хордомезодермального кармана (архентерона) с субгерминативной полостью (образование, аналогичное нервно-кишечному каналу). При этом закладка хорды по средней линии включается в свод энтодермальной закладки кишечника. В эктодерме каудально от первичной пластинки образуется утолщенная первичная полоска, и из ее области, так же как и у птиц, материал парахордральной мезодермы, располагающийся по обеим сторонам хорды, перемещается во внутрь. И у птиц после окончания гаструляции и образования энтодермы в области эмбриональной зоны (в зародышевом щитке) начинается нотогенез.
Сущность гаструляции заключается в том, что однослойный зародыш — бластула — превращается в многослойный — двух- или трехслойный, называемый гаструлой (греч., gaster — желудок; позднелат., gastrula – желудочек).
У двухслойных примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок — эктодерму — и внутренний зародышевый листок — энтодерму. Энтодерма формирует первичную кишку с полостью внутри — гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Два зародышевых листка являются определяющим морфологическим признаком гаструляции. Их существование на определенной стадии развития у всех многоклеточных животных, начиная с кишечнополостных и кончая высшими позвоночными, позволяет думать о гомологии зародышевых листков и единстве происхождения всех этих животных.
У позвоночных помимо двух упомянутых зародышевых листков во время гаструляции образуется еще третий — мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка можно расценивать как эволюционное усложнение стадии гаструляции у позвоночных, связанное с ускорением у них развития в ранних фазах эмбриогенеза. У примитивных хордовых животных, таких, как ланцетник, аналог 3-го зародышевого листка хордомезодерма обычно образуется в начале следующей после гаструляции стадии — первичного органогенеза. Изменение времени закладки органов и структур в процессе эволюции (гетерохрония) встречается достаточно часто.
Процесс гаструляции характеризуется важными клеточными преобразованиями, такими, как направленные перемещения клеточных групп (пластов) и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий. Перечисленные клеточные механизмы онтогенеза – см. 8.2.
Способы гаструляции различны. Выделяют четыре разновидности направленных перемещений клеток, приводящих к преобразованию зародыша из однослойного в многослойный.
Инвагинация — впячивание одного из участков бластодермы внутрь целым пластом. У ланцетника впячиваются клетки вегетативного полюса, у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсами в области серого серпа (см. рис. 7-3). Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка.
Эпиболия — обрастание мелкими быстро делящимися клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных.
Деламинация— расслоение клеток бластодермы на два слоя, лежащих друг над другом. Деламинацию можно наблюдать в дискобластуле зародышей с неполным дроблением, таких, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деламинация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта.
Иммиграция — перемещение клеточных групп или отдельных клеток, не объединенных в единый пласт. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных (рептилии, птицы, млекопитающие).
Дата добавления: 2016-05-05; просмотров: 2040;