А 2. Физиология центральной нервной системы
2.1.1. Структурнофункциональная характеристика центральной нервной системы(рис. 27).
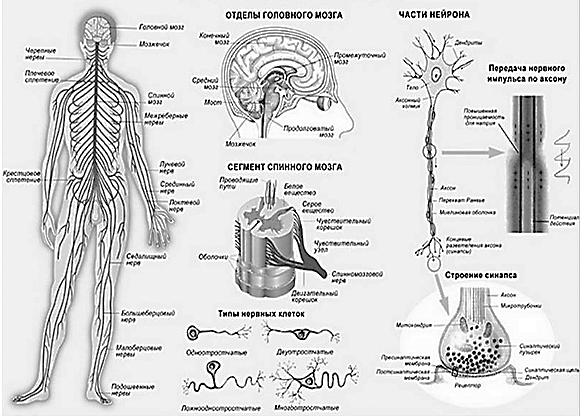
Рис. 27. Основные структурные элементы нервной системы. Слева: общий план строения нервной системы. Посередине: основные структурные элементы спинного и голвного мозга, типы нейронов. Справа: структурные части нейрона и синапса
Нервная клетка (нейрон) является структурной и функциональной единицей ЦНС. Она состоит из сомы (тела клетки с ядром) и отростков, представляющих собой большое число дендритов и один аксон (рис. 28).
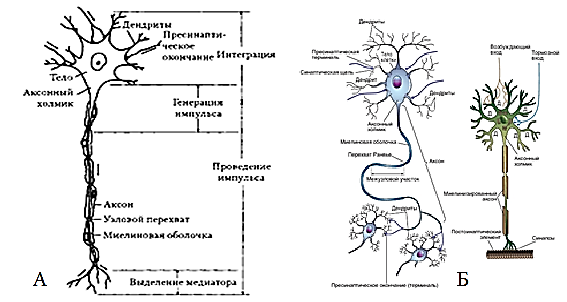
Рис. 28. Структурно-функциональные элементы мотонейрона (А) спинного мозга (Агаджанян, 2007) и синаптические контакты нейрона – тормозной и возбуждающий синапсы (Б) (Камкин, 2010)
Любая деятельность самой ЦНС реализуется в конечном итоге с помощью функционирования отдельных клеток. ЦНС человека содержит свыше 100 млрд нервных клеток, взаимодействие между которыми осуществляется посредством множества синапсов. Число последних в тысячи раз больше количества самих клеток (1015- 1016), так как их аксоны многократно делятся дихотомически. Нейроны оказывают влияние на органы и ткани также посредством синапсов. Важную роль в деятельности нервной системы играют глиальные клетки и ликвор. Развитие неврологических нарушений у больных СПИДом связано с гибелью 40—50% корковых нейронов. Нервные клетки имеются и вне ЦНС: нейроны ганглиев ВНС, афферентные нейроны спинномозговых ганглиев и ганглиев черепных нервов. Однако периферических нервных клеток, по сравнению с центральными, мало — всего лишь около 25 млн.
В теле нейрона синтезируются макромолекулы, которые транспортируются по аксону и дендритам; оно выполняет трофическую функцию по отношению к своим отросткам и клеткам-эффекторам. Отросток, лишенный связи с сомой нейрона, дегенерирует. ПП нейрона составляет —60—80 мВ, ПД — 80—110 мВ. Скопления тел нейронов образуют серое вещество мозга, отростков — белое вещество. Сома и дендриты каждого нейрона покрыты нервными окончаниями — синаптическими бутонами других нервных клеток и отростками глиальных клеток. На одном мотонейроне число синаптических бутонов может достигать 10—20 тыс. (см. рис. 27).
Аксон начинается от тела клетки аксонным холмиком. Диаметр тела клетки составляет 10—100 мкм, аксона — 1—6 мкм; на периферии длина аксона может достигать 1 м и более. По аксону передаются нервные импульсы к другим нейронам и эффекторным клеткам.
Нейрон имеет большое число дендритов, на которых локализуется большинство синапсов нейрона, обеспечивающих (вместе с синапсами сомы) восприятие информации от других нервных клеток.
Основные функции нейрона — восприятие, переработка и хранение информации, передача сигнала другим нервным клеткам, регуляция деятельности клеток различных органов и тканей организма.
Отдельные структурные элементы нейрона выполняют определенные функции.
Структурами, воспринимающими импульсы от других нервных клеток, являются тело и дендриты нейрона с расположенными на них шипиками. Если шипики не получают импульсацию, то они исчезают. Импульсы могут поступать и к окончанию аксона — аксоаксонные синапсы, например в случае пресинаптического торможения. Число дендритов у нейрона весьма вариабельно — от 1 до 1500.
Структура, где начинает формироваться ПД, — аксонный холмик.
Структура, проводящая возбуждение к другому нейрону или к эффекторной клетке, — аксон.
Структура, передающая импульсы на другую клетку, — это пресинаптический аппарат, представляющий собой нервное окончание с пресинаптической мембраной. В нем синтезируется и запасается медиатор. При возбуждении нервного окончания медиатор выбрасывается в синаптическую щель через пресинаптическую мембрану, которая обратно захватывает неиспользованный медиатор в нервное окончание.
В процессе деятельности нервной системы отдельные нейроны образуют нейронные цепи (например, рефлекторная дуга), объединяются в ансамбли (модули), нейронные сети. Последние могут включать несколько нейронов, десятки, тысячи нейронов, при этом совокупность клеток, образующих модуль, обеспечивает появление новых свойств, которыми не обладают отдельные нейроны. Деятельность каждой клетки в составе модуля становится функцией не только поступающих к ней сигналов, но и функцией процессов, обусловленных той или иной конструкцией модуля (П. Г. Костюк)! Примером восходящих сетей являются сенсорные афферентные пути.
Глиальные клетки (нейроглия — «нервный клей»; от греч. neuron — нерв, glia — клей) более многочисленны, чем нейроны, составляют около 50% объема ЦНС; способны к делению в течение всей жизни. По размерам глиальные клетки в 3—4 раза меньше нервных. Число их огромно — 14—1010 (в 10 раз больше, чем нейронов), с возрастом оно увеличивается (число нейронов уменьшается). Глиальные клетки окружают тела нейронов и их аксоны. Мембранный потенциал клеток нейроглии составляет -70 —90 мВ. Электрические поля их ЛП взаимодействуют с электрическими полями ВПСП и ПД нейронов.
Глиальные клетки выполняют несколько функций: изолирующую (глиоциты окутывают нейроциты в виде своеобразного футляра и продуцируют миелин), опорную, защитную (выработка цитокинов, фагоцитоз, при повреждении нейронов образуют глиальный рубец), обменную (синтез белков памяти, снабжение нейронов питательными веществами, поддержание оптимальной концентрации ионов К+ в интерстиции). Микроглиальные клетки способны к ритмическому изменению своего объема (период «сокращения»— 1,5 мин, «расслабления» — 4 мин). Циклы изменения объема повторяются через каждые 2—20 ч. Полагают, что пульсация способствует продвижению аксоплазмы в нейронах и влияет на ток межклеточной жидкости. Норадреналин усиливает, серотонин угнетает пульсацию глиоцитов. Глиоциты, охватывая снаружи капилляры ЦНС, участвуют в формировании ГЭБ.
Ликвор — бесцветная прозрачная жидкость, заполняющая мозговые желудочки, спинномозговой канал и субарахноидальное пространство (Рис. 29).
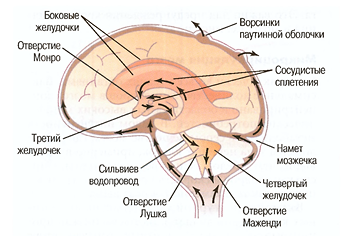
Рис. 29. Стрелками показан путь спинномозговой жидкости от сосудистых сплетений боковых желудочков к ворсинкам паутиной оболочки, выступающим в синусы твердой мозговой оболочки (Гайтон, 2008)
Происхождение ликвора связано с интерстициальной жидкостью мозга, значительная часть его (ликвора) образуется сосудистыми сплетениями желудочков мозга — это совокупность фильтрата плазмы крови и интерстициальной жидкости. От плазмы крови ликвор отличается, как и межклеточная жидкость других тканей, низким содержанием белка (0,1 г/л, а в плазме крови 75 г/л), меньшим содержанием аминокислот (0,8 и 2,0 ммоль/л соответственно) и глюкозы (3,9 и около 5,0 ммоль/л соответственно). Его объем 100-200 мл (12—14% общего объема мозга), за сутки вырабатывается около 600 мл. Обновление этой жидкости происходит 4-8 раз в сутки; давление ликвора при горизонтальном положении человека составляет 7—14 мм рт. ст., при вертикальном — 15—30 мм рт. ст.
Функции ликвора: выполняет защитную роль (она обладает бактерицидными свойствами, содержит иммуноглобулины G и А, систему комплемента, моноциты и лимфоциты, является своеобразной гидравлической «подушкой» мозга).
Интерстициальная жидкость является непосредственной питательной средой клеток мозга, в нее клетки выделяют также и продукты своего обмена.
Отток ликвора выполняет дренажную функцию (препятствие оттоку ликвора в кровь может сопровождаться тяжелыми нарушениями в деятельности ЦНС). Отток ликвора осуществляется несколькими путями: 30—40% — через субарахноидальное пространство в продольный синус венозной системы головного мозга; 10—20% — через периневральные пространства черепных и спинномозговых нервов в лимфатическую систему; часть ликвора реабсорбируется сосудистыми сплетениями мозга.
Нейроны ЦНС классифицируют по нескольким признакам.
По влиянию на другие клетки имеются возбуждающиеи тормозныенейроны.
По виду медиатора, выделяющегося в окончания аксонов, различают нейроны серотонинергические, адренергические, холинергические и др.
По специфичности воспринимаемой сенсорной информации нейроны высших отделов ЦНС могут быть моно-, би- и полисенсорными. Например, моносенсорными являются нейроны первичных сенсорных зон в коре большого мозга. Первичные афферентные нейроны (спинальных ганглиев и ганглиев черепных нервов) также моносенсорны: они воспринимают с помощью собственных рецепторов только один вид раздражителя (адекватного) — свет, химическое вещество, электрическое, механическое и температурное воздействие. Бисенсорные нейроны встречаются во вторичных сенсорных зонах в коре большого мозга. Полисенсорные нейроны — это нейроны ассоциативных зон мозга (третичные сенсорные зоны), моторной коры, они реагируют на любые раздражители (звук, свет, прикосйовение и др.).
По активности имеются фоновоактивные и «молчащие» нейроны, возбуждающиеся только в ответ на раздражение. Фоновоактивные нейроны играют важную роль в поддержании тонуса ЦНС и, особенно, коры большого мозга. Фоновоактивные нейроны различаются по общему рисунку генерации импульсов, так как одни нейроны разряжаются непрерывно (ритмично или аритмично), другие — «пачками» импульсов.
По направлению передачи информации нейроны подразделяют на: 1) афферентные, воспринимающие с помощью рецепторов информацию о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; 2) эфферентные, передающие информацию к рабочим органам — эффекторам; нервные клетки, иннервирующие эффекторы, иногда называют эффекторными (эффекторные нейроны спинного мозга — мотонейроны делят на α- и γ-мотонейроны); 3) вставочные (интернейроны), обеспечивающие взаимодействие между нейронами ЦНС различных уровней.
В зависимости от отдела нервной системы выделяют нейроны соматической и вегетативной нервной системы.
Синапсы ЦНС классифицируют также по нескольким признакам.
По эффекту, как и нейроны, — возбуждающие и тормозные.
В зависимости от местоположения — аксосоматические, аксодендритные, аксо-аксонные, дендро-дендритные, дендросоматические.
По механизму передачи сигнала к другим клеткам: 1) химические синапсы (наиболее распространенный способ); 2) электрические синапсы (редко); 3) смешанные синапсы (электрохимические), которые обнаружены между нейронами латерального вестибулярного ядра; 4) часть нейронов гипоталамуса информацию передают с помощью гормонов.
Химические синапсы имеют относительно широкую синаптическую щель (20-50 нм). В пресинаптической терминали содержится большое число пузырьков — пресинаптических везикул (диаметр около 50 нм), заполненных медиатором — химическим посредником. Медиатор либо образуется в теле нейрона и транспортируется в пресинаптическое окончание аксонным транспортом, либо синтезируется непосредственно в пресинаптическом окончании и накапливается в везикулах.
Для синтеза медиатора нужны ферменты, образующиеся в теле клетки на рибосомах. Медиатор в пресинаптическом окончании находится в двух пулах— готовом к экзоцитозу (пул 1, медиатор лежит на пресинаптической мембране изнутри) и мобилизационном (пул 2, находится на цитоскелете), из которого пополняется в течение нескольких секунд пул 1. Этот процесс активируется ионами Са2+.
Химические синапсы передают сигнал относительно медленно, односторонне, менее надежно, чем электрические синапсы. Химическим синапсам присуще явление суммации. Электросекреторное сопряжение в синапсах ЦНС, как и в нервно-мышечных синапсах, обеспечивается ионами Са2+ — при возбуждении пресинаптической терминали открываются потенциалуправляемые Са-каналы, дополнительно Са2+ выделяется из эндоплазматического ретикулума. Кальций входит в нервное окончание согласно электрохимическому градиенту и запускает процесс высвобождения медиатора в синаптическую щель с помощью экзоцитоза — ион Са2+ активирует белковый экзоцитозный аппарат пресинапса, представляющий собой совокупность белков (синапсин, спектрин), активация которых обеспечивает выход медиатора в синаптическую щель.
Синаптическая щель химических синапсов содержит поперечно расположенные гликопротеидные филаменты, обеспечивающие связи пре- и постсинаптической мембран и направление диффундирующего медиатора от пресинаптической мембраны к постсинаптической. Выделившийся в синаптическую щель медиатор взаимодействует с белком-рецептором постсинаптической мембраны, что и приводит нервную клетку в состояние возбуждения или торможения.
Электрические синапсы имеются возбуждающие и тормозные (последних мало, изучены они недостаточно). Электрические синапсы имеют щель на порядок меньшую (2-4 нм), чем у химических синапсов, проводят сигнал в обе стороны без синаптической задержки, передача не блокируется при удалении Са2+; они малочувствительны к фармакологическим препаратам и ядам, практически не утомляемы, как и нервное волокно. Контактирующие мембраны клеток в составе электрического синапса связаны друг с другом полуканалами белковой природы — коннексонами. Очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран обеспечивает хорошую электрическую проводимость — за счет электрического поля. Электрические возбуждающие синапсы обнаружены в головном мозге млекопитающих в составе мезэнцефального ядра тройничного нерва, вестибулярного ядра Дейтерса, ядра нижней оливы продолговатого мозга. Имеются следующие электрические синапсы: аксон—сома; аксон—дендрит; аксон—аксон; дендрит—дендрит; дендрит—сома; сома—сома. Электрические синапсы характерны для сердца, гладких мышц, секреторных клеток. Через коннексоны нейроны ЦНС обмениваются некоторыми компонентами цитоплазмы — пептидами, РНК, метаболитами, циклическими нуклеотидами, аминокислотами. Электрические синапсы оказывают влияние на метаболизм контактирующих клеток.
Работа электрических синапсов может регулироваться близлежащими химическими синапсами.
По механизму активации постсинаптические рецепторы химических синапсов делятся на два класса: ионотропные и метаботропные (Рис. 30).
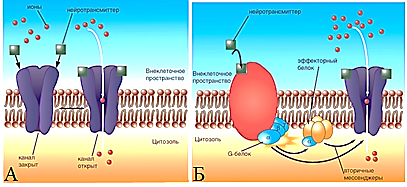
Рис. 30. Ионотропные и метаботропные рецепторы: А – ионотропный рецептор, Б – метаботропный рецептор (см. описание в тексте) (Камкин, 2010)
Ионотропные, активация ионных каналов которых осуществляется нейромедиатором непосредственно с помощью G-белка (например, N-холинорецептор);
Метаботропные (например, α1- и α2-адренорецепторы), когда ионные каналы активируются через G-белок с помощью вторых посредников (цАМФ, цГМФ, ИФ3, ДАГ). После каждого цикла передачи сигнала выделившийся медиатор инактивируется специфическим ферментом (например, ацетилхолин — ацетилхолинэстеразой, норадреналин — моноаминоксидазой (МАО), обратно захватывается пресинапсом, часть медиатора рассеивается.
Функции ЦНС. Нервная система выполняет интегрирующую роль. Она объединяет в единое целое все ткани, органы организма, координируя их специфическую активность в составе целостных гомеостатических и поведенческих функциональных систем.
Основные частные функции нервной системы:
1. Обеспечение, сознания и всех видов психической деятельности.
2.Управление деятельностью опорно-двигательного аппарата.
3. Нервная система регулирует тонус мышц и посредством его перераспределения поддерживает естественную позу, а при нарушении последней восстанавливает ее; инициирует все виды движений в процессе трудовой деятельности, занятий физкультурой и спортом.
3.Регуляция работы внутренних органов, осуществляемая с помощью ВНС и эндокринных желез. При этом в покое обеспечивается гомеостазис (постоянство внутренней среды организма), а во время работы — приспособительное регулирование деятельности внутренних органов согласно потребностям организма и поддержание гомеостазиса.
4.Формирование целенаправленного поведения организма в социальном обществе и окружающей среде. Оно реализуется с помощью реакций избегания или избавления от неприятных раздражителей (защитные реакции организма), регуляции интенсивности обмена веществ при изменении температуры окружающей среды. Изменения внутренней среды организма, воспринимаемые субъективно в виде ощущений (жажда, голод), также побуждают организм к той или иной целенаправленной деятельности.
В таблице 4 кратко представлены функции нервной системы.
Таблица 4. Отделы центральной нервной системы и их функции (Камкин, 2008)
| Отдел нервной системы | Функция |
| Спинной мозг | Сенсорный вход, рефлексы: соматический и вегетативный (автономный), двигательный выход |
| Продолговатый мозг | Регуляция сердечно-сосудистых функций и дыхания; рефлексы ствола мозга |
| Мост | Регуляция дыхания и мочеиспускания; вестибулярная регуляция движений глаз |
| Мозжечок | Регуляция движений; двигательное научение |
| Средний мозг | Переключение слуховых путей; регуляция движений глаз; регуляция движений |
| Таламус | Переключение сенсорных путей к коре |
| Гипоталамус | Регуляция вегетативных и эндокринных функций |
| Базальные ганглии (ядра) | Регуляция движений |
| Лимбическая система | Участие в процессах памяти; в эмоциях и инстинктах, регуляция вегетативных функций (через гипоталамус) |
| Кора больших полушарий | Сенсорное восприятие; когнитивные функции; обучаемость и память; планирование движений и произвольные движения |
Дата добавления: 2016-02-16; просмотров: 4441;