Механизм сокращения скелетной мышцы
Укорочение мышцы является результатом сокращения множества саркомеров. При укорочении актиновые нити скользят относительно миозиновых, в результате чего длина каждого саркомера мышечного волокна уменьшается. При этом длина самих нитей остается неизменной. Миозиновые нити имеют поперечные выступы (поперечные мостики) длиной около 20 нм. Каждый выступ состоит из головки, которая соединена с миозиновой нитью посредством «шейки» (рис. 23).
При расслабленном состоянии мышцы головки поперечных мостиков не могут взаимодействовать с актиновыми нитями, поскольку их активные участки (места взаимного контакта с головками) изолированы тропомиозином. Укорочение мышцы является результатом конформационных изменений поперечного мостика: его головка совершает наклон с помощью сгибания «шейки».
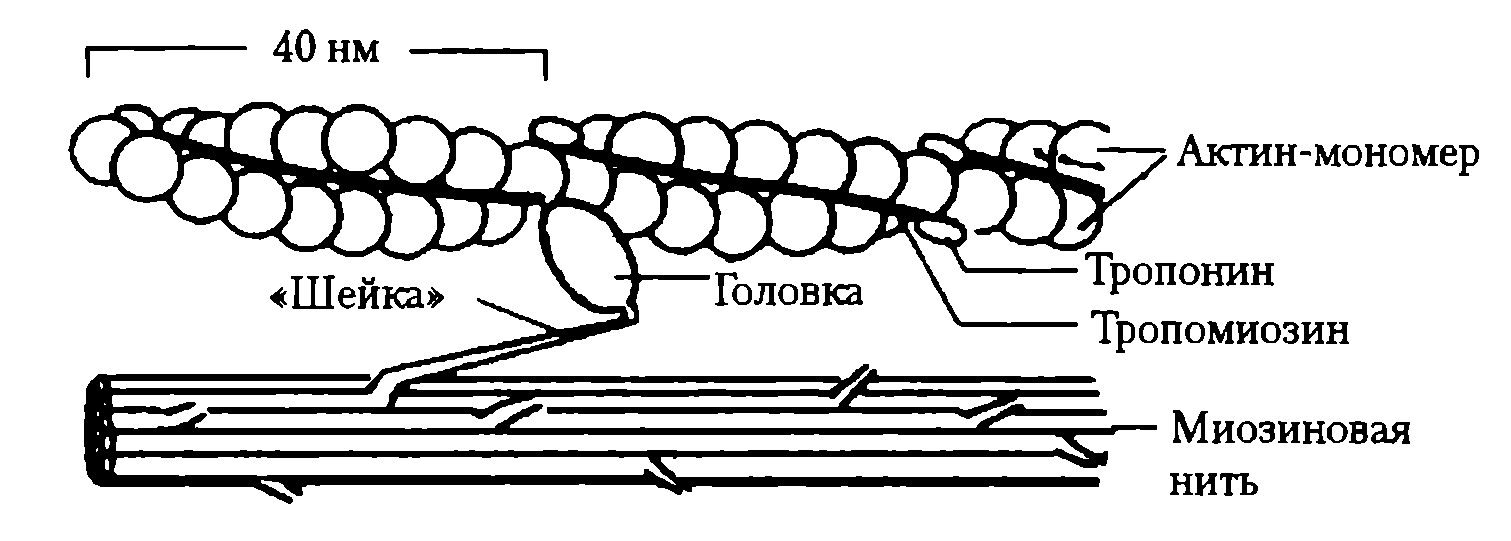
Рис. 23. Ространственная организация сократительных и регуляторных белков в исчерченной мышце. Показано положение миозинового мостика (гребковый эффект, шейка согнута) в процессе взаимодействия сократительных белков в мышечных волокне (сокращение волокна)
Последовательность процессов, обеспечивающих сокращение мышечного волокна (электромеханическое сопряжение):
1. После возникновения ПД в мышечном волокне вблизи синапса (за счет электрического поля ПКП) возбуждение распространяется по мембране миоцита, в том числе по мембранам поперечных Т-трубочек. Механизм проведения ПД по мышечному волокну такой же, как и по безмиелиновому нервному волокну — возникший ПД вблизи синапса посредством своего электрического поля обеспечивает возникновение новых ПД в соседнем участке волокна и т.д. (непрерывное проведение возбуждения).
2. Потенциал действия Т-трубочек за счет своего электрического поля активирует потенциалуправляемые кальциевые каналы на мембране СПР, вследствие чего Са2+ выходит из цистерн СПР согласно электрохимическому градиенту.
3. В межфибриллярном пространстве Са2+ контактирует с тропонином, что приводит к его конформации и смещению тропомиозина, в результате чего на нитях актина обнажаются активные участки, с которыми соединяются головки миозиновых мостиков.
4. В результате взаимодействия с актином АТФазная активность головок миозиновых нитей усиливается, обеспечивая освобождение энергии АТФ, которая расходуется на сгибание миозинового мостика, внешне напоминающего движение весел при гребле (гребковое движение) (см. рис. 23), обеспечивающее скольжение актиновых нитей относительно миозиновых. На совершение одного гребкового движения расходуется энергия одной молекулы АТФ. При этом нити сократительных белков смещаются на 20 нм. Присоединение новой молекулы АТФ к другому участку головки миозина ведет к прекращению зацепления ее, но при этом энергия АТФ не расходуется. При отсутствии АТФ головки миозина не могут оторваться от актина — мышца напряжена; таков, в частности, механизм трупного окоченения.
5. После этого головки поперечных мостиков в силу своей эластичности возвращаются в исходное положение и устанавливают контакт со следующим участком актина; далее вновь происходит очередное гребковое движение и скольжение актиновых и миозиновых нитей. Подобные элементарные акты многократно повторяются. Одно гребковое движение (один шаг) вызывает уменьшение длины каждого саркомера на 1%. При сокращении изолированной мышцы лягушки без нагрузки 50% укорочение саркомеров происходит за 0,1 с. Для этого необходимо совершение 50 гребковых движений. Миозиновые мостики сгибаются асинхронно, но в связи с тем, что их много и каждая миозиновая нить окружена несколькими актиновыми нитями, сокращение мышцы происходит плавно.
Расслабление мышцы происходит благодаря процессам, протекающим в обратной последовательности. Реполяризация сарколеммы и Т-трубочек ведет к закрытию кальциевых потенциалуправляемых каналов мембраны СПР. Са-насосы возвращают Са2+ в СПР (активность насосов возрастает при увеличении концентрации свободных ионов).
Снижение концентрации Са2+ в межфибриллярном пространстве вызывает обратную конформацию тропонина, в результате чего тропомиозиновые нити изолируют активные участки актиновых филаментов, что делает невозможным взаимодействие с ними головок поперечных мостиков миозина. Скольжение актиновых нитей вдоль миозиновых в обратном направлении происходит под действием сил гравитации и эластической тяги элементов мышечного волокна, что восстанавливает исходные размеры саркомеров.
Источником энергии для обеспечения работы скелетных мышц является АТФ, расходы которой значительны. Даже в условиях основного обмена на функционирование мускулатуры организм затрагивает около 25% всех своих энергоресурсов. Затраты энергии резко возрастают во время выполнения физической работы.
Запасы АТФ в мышечном волокне незначительны (5 ммоль/л) и могут обеспечить не более 10 одиночных сокращений.
Расход энергии АТФ необходим для осуществления следующих процессов.
Во-первых, энергия АТФ расходуется на обеспечение работы Nа/К-насоса (он поддерживает градиент концентрации Na+ и К+ внутри и вне клетки, формирующих ПП и ПД, обеспечивающего электромеханическое сопряжение) и работы Са-насоса, который понижает концентрацию Са2+ в саркоплазме после сокращения мышечного волокна, что приводит к расслаблению.
Во-вторых, энергия АТФ расходуется на гребковое движение миозиновых мостиков (сгибание их).
Ресинтез АТФ осуществляется с помощью трех энергетических систем организма.
1. Фосфогенная энергетическая система обеспечивает ресинтез АТФ за счет имеющегося в мышцах высокоэнергоемкого КФ и образовавшейся при расщеплении АТФ аденозиндифосфорной кислоты (аденозиндифосфат, АДФ) с образованием креатина (К): АДФ + + КФ → АТФ + К. Это мгновенный ресинтез АТФ, при этом мышца может развивать большую мощность, но кратковременно — до 6 с, поскольку запасы КФ в мышце ограниченны.
2. Анаэробная гликолитическая энергетическая система обеспечивает ресинтез АТФ за счет энергии анаэробного расщепления глюкозы до молочной кислоты. Этот путь ресинтеза АТФ является быстрым, но тоже кратковременным (1—2 мин), так как накопление молочной кислоты тормозит активность гликолитических ферментов. Однако лактат, вызывая местный сосудорасширяющий эффект, улучшает кровоток в работающей мышце и снабжение ее кислородом и питательными веществами.
3. Аэробная энергетическая система обеспечивает ресинтез АТФ с помощью окислительного фосфорилирования углеводов и жирных кислот, протекающего в митохондриях мышечных клеток. Этот способ может обеспечить энергией работу мышц в течение нескольких часов и является основным способом энергетического обеспечения работы скелетных мышц.
Дата добавления: 2016-02-16; просмотров: 4449;