Физиология нервных волокон
Структурно-функциональная характеристика нервных волокон. Нервные волокна — это отростки нейронов, с помощью которых осуществляется связь между нейронами и с исполнительными клетками. Имеется два типа нервных волокон: миелинизированные и немиелинизированные (безмиелиновые). Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты), в которые погружаются осевые цилиндры нервных волокон. Оболочку миелинизированных волокон образуют в периферической нервной системе также шванновские клетки (миелоциты, глиальная клетка), формирующие миелин (многослойная обертка мембран миелоцитов — до 100), а в ЦНС — олигодендроциты (глиальные клетки, т.е. те же шванновские клетки). Миелиновая оболочка через равные участки (около 1 мм) прерывается, образуя свободные от миелина небольшие участки — перехваты Ранвье. Основную часть миелина (78% сухого веса) составляют липиды, обеспечивающие изолирующие свойства оболочки. Нервные волокна обеспечивают проведение возбуждения и аксонныйтра транспорт, выполняющий трофическую функцию нейрона.
Выделяют три типа волокон: А, В и С (см. табл. 3).
Таблица 3. Типы волокон в нервах млекопитающих (по Эрлангеру-Гассеру)
| Тип волокон | Диаметр волокна, мкм | Скорость проведения возбуждения, м/с | Длительность абсолютного рефрактерного периода, мс |
| Аα (миел.) | 12-20 | 70-120 | 0,4-1,0 |
| Аβ (миел.) | 5-12 | 30-70 | - |
| Аσ (миел.) | 3-6 | 15-30 | 0.4-1,0 |
| Аγ (миел.) | 2-5 | 12-30 | - |
| В (миел.) | 1-3 | 5-12 | 1,2 |
| С (немиел.) | 0,3-1,3 | 0,5-2,3 |
Волокна типов А и В являются миелинизированными. А-волокна — это афферентные и эфферентные волокна соматической нервной системы; к волокнам типа В принадлежат преганглионарные волокна ВНС. С-волокна немиелинизированные — это постганглионарные волокна ВНС, а также афферентные волокна от некоторых болевых, тепловых и висцеральных рецепторов.
Аксонный транспорт. Основная масса веществ образуется в трофическом центре нейрона, расположенном преимущественно около ядра, а используются эти вещества в различных участках тела нейрона и его отростках. В аксонных окончаниях также происходит синтез медиаторов, АТФ и повторное использование мембраны пузырьков после высвобождение медиатора. Выделяют быстрый и медленный аксонный транспорт (оба они с затратой энергии).
Быстрый аксонный транспорт осуществляется со скоростью 200—400 мм/сут от тела клетки до аксонных окончаний — прямой (антероградный) — и в противоположном направлении — обратный (ретроградный) транспорт. Вещества транспортируются с помощью микротрубочек и микрофиламентов, часть которых представляет собой актиновые нити (актин составляет 10—15 % белков нейрона). Посредством прямого транспорта в аксонные окончания доставляются митохондрии, ферменты, медиаторы, липиды, везикулы, содержащие гликопротеиды мембран, специальные белки и пептиды (нейротрофогены). Посредством обратного транспорта в тело нейрона переносятся везикулы, содержащие остатки разрушенных структур, фрагменты мембран, факторы роста нервов и другие ростовые факторы, регулирующие синтез белка в соме клетки. Многие вещества, доставленные посредством ретроградного транспорта, подвергаются разрушению в лизосомах. В патологических случаях по аксону к телу клетки могут транспортироваться столбнячный экзотоксин, вирусы полиомиелита, герпеса, бешенства.
Медленный аксонный транспорт идет в прямом направлении и представляет собой передвижение всего столба аксоплазмы со скоростью 1—2 мм/сут. С помощью этого транспорта перемещаются образованные в эндоплазматической сети белки микротрубочек и микрофиламентов (актин, тубулин и др.), ферменты цитозоля, РНК, белки каналов, насосов и другие вещества.
Значение аксонного транспорта: 1) необходим для поддержания структуры нервного волокна; 2) необходим для аксонного роста и образования синаптических контактов; 3) играет важную роль при регенерации нервных волокон. На мышечное волокно такое влияние оказывают нейротрофогены (специальные белки, β-эндорфин и другие пептиды); обратное влияние на мотонейрон осуществляется с помощью миотрофогенов (фактора роста нервов, инсулиноподобного фактора роста).
Результат повреждения нервного волокна. Если нервное волокно в результате травмы разорвано, его периферический отрезок, лишенный связи с телом нейрона, подвергается разрушению, которое называется валлеровской дегенерацией. В течение 2-3 сут наступает распад нейрофибрилл, митохондрий, миелина и синаптических окончаний. Участок волокна, связанный с телом нейрона, регенерирует, проводимость восстанавливается. В норме нервное волокно, как и нейрон, функционирует на протяжении всей жизни организма и проводит возбуждение без декремента (затухания) по всей длине волокна, например от тела мотонейронов спинного мозга до мышечных волокон конечностей (до 1 м).
Механизм проведения возбуждения. Проведение ПД возможно только при наличии на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование новых ПД. Это осуществляется, согласно нашим представлениям, следующим образом. В распространении ПД можно выделить два этапа: этап распространения электрического поля, снижающего мембранный потенциал, и этап генерации новых ПД в новых участках нервного волокна. Электрическое поле — разновидность материи, посредством которой осуществляется силовое воздействие на электрические заряды, находящиеся в этом поле. Электрическое поле, которое генерируется биологическими структурами, является источником информации о состоянии клеток и органов организма. Например, состояние электрического поля сердца, записанного в виде электрокардиограммы, помогает выявить его возможные повреждения. В зависимости от расположения и концентрации ионных каналов, в мембране нервного или мышечного волокна имеется два варианта проведения ПД: непрерывное и сальтаторное (скачкообразное).
Непрерывное проведение ПД происходит в мышечных волокнах и в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалуправляемых ионных каналов по всей длине волокна. Проведение нервного импульса начинается (как и в мышечном волокне) с распространения колеблющегося по величине электрического поля. Амплитуда ПД в нервном волокне (мембранный потенциал + инверсия) составляет 100-120 мВ, постоянная длины мембраны (λm— расстояние, на котором сохраняется 37 % величины ПД в виде электрического поля) в безмиелиновых волокнах равна 0,1—1,0 мм. В связи с этим возникший ПД за счет действия своего электрического поля способен деполяризовать мембрану соседнего участка до критического уровня на расстояние от 0,1 до 1,0 мм. Это означает, что на этом участке (0,1-1,0 мм) одновременно генерируются новые ПД, обусловленные движением ионов Na+ в клетку, К+— из клетки (на распространение электрического поля время не затрачивается). Число одновременно возникающих ПД ограничивается длиной возбужденного участка — для безмиелинового волокна 0,1-1,0 мм (ПД возникают рядом друг с другом в непосредственной близости). Причем сами ПД не перемещаются (они исчезают там, где возникают). Главную роль в возникновении новых ПД играет передний ПД. Вспомогательную роль в генерации новых ПД в невозбужденных участках нервного волокна играют промежуточные ПД (возникшие сзади переднего ПД) — их электрическое поле суммируется с электрическим полем переднего ПД, но они находятся дальше от участка нервного волокна, где возникают новые ПД. Таким образом, непрерывное распространение нервного импульса, как и сальтаторное (см. ниже), идет через генерацию новых ПД по эстафете, когда каждый участок мембраны сначала выступает как раздражаемый электрическим полем, а затем — как раздражающий (в результате формирования в нем новых ПД) (рис. 15).
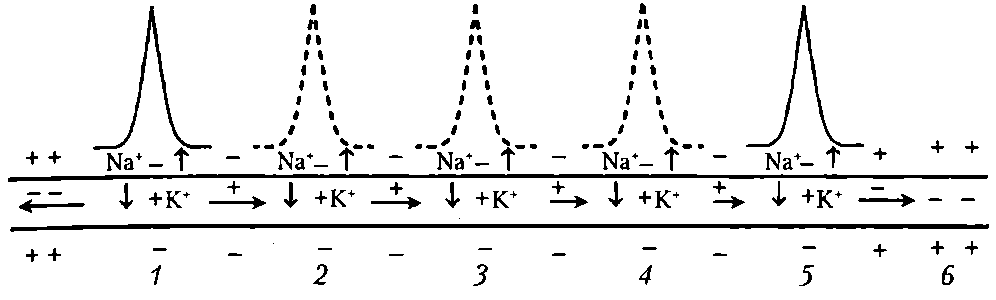
Рис. 15. Непрерывное проведение возбуждения (ПД) в безмиелиновом нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1-5 - состояние возбуждения (ПД); 6 – состояние покоя; пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na+ в клетку и K+ - из клетки
Сальтаторное проведение ПД происходит по миелинизированным волокнам (типы А и В), для которых характерна концентрация потенциалуправляемых ионных каналов только в небольших участках мембраны (в перехватах Ранвье), где их плотность достигает 10 тыс. на 1 мкм2, что примерно в 100 раз выше, чем в любом участке мембраны безмиелиновых волокон. В области миелиновых муфт, обладающих хорошими изолирующими свойствами, потенциалуправляемых каналов почти нет, поэтому ПД здесь не возникают. ПД, возникший в одном перехвате Ранвье, за счет действия своего электрического поля деполяризует мембрану соседних перехватов до КП, что приводит к возникновению в них новых ПД, т.е. возбуждение возникает как бы скачкообразно — только в перехватах (рис.16).
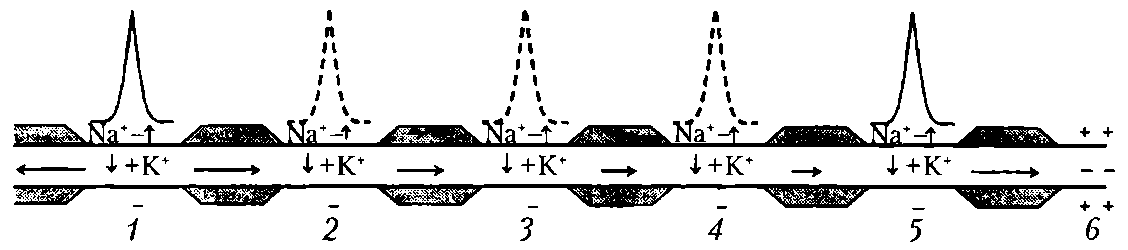
Рис. 16. Сальтаторное проведение возбуждения (ПД) в миелинизированном нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1-5 - состояние возбуждения (ПД); 6 – состояние покоя; пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na+ в клетку и K+ - из клетки
Напомним, что Nа-каналы начинают открываться при достижении деполяризации клеточной мембраны 50% КП. Постоянная длины мембраны миелинового волокна достигает 5 мм. Это означает, что электрическое поле ПД на данном расстоянии сохраняет 37% своей амплитуды (около 30 мВ) и может деполяризовать мембрану до КП (ΔV в перехватах Ранвье составляет около 15 мВ). Благодаря этому, в случае повреждения ближайших на пути следования перехватов Ранвье ПД возбуждает 2—4-й и даже 5-й перехваты. Поэтому возбуждение распространяется очень быстро по всей длине волокна, а ионы движутся только перпендикулярно относительно длины волокна — в клетку и из клетки (вдоль волокна они не успевают смещаться). Электрическое поле ПД, возникших сзади переднего (промежуточные ПД), суммируется с электрическим полем переднего ПД, как и при непрерывном распространении возбуждения.
Простые расчеты показывают, что скорость проведения ПД по нервному волокну за счет движения ионов вдоль волокна была бы слишком низкой. В частности, скорость движения Na+ в клетку согласно электрохимическому градиенту легко рассчитать по толщине клеточной мембраны (6—10 нм) и длительности ПД (например, толстого миелинизированного волокна — около 1 мс) — вход Na+ в клетку и выход К+ из клетки. При этом Na+ при движении в клетку согласно электрохимическому градиенту преодолевает расстояние в 8 нм примерно за 0,5 мс (восходящая часть пика ПД продолжается около 0,5 мс). На основании этого рассчитаем, сколько времени потребуется на прохождение ПД 1 м. Для прохождения ионом Na+ 8 нм необходимо 0,5 мс.
Отсюда:
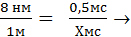
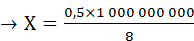 ≈17 часов,
≈17 часов,
т.е. возбуждение распространилось бы на 1 мм за 17 часов.
Легко представить, какова была бы подвижность (точнее, неподвижность) живых существ, если бы возбуждение вдоль нервных и мышечных волокон распространялось со скоростью диффузии ионов, но очень трудно представить, как бы выглядел весь животный мир! Чтобы шевельнуть пальцем пришлось бы ждать около суток!
Возникающие ПД не могут инициировать развитие других ПД в обратном направлении, так как нервное волокно находится еще в рефрактерном состоянии. Это не противоречит тому факту, что раздражение нервного волокна в эксперименте вызывает распространение возбуждения в двух направлениях, поскольку в этом случае участки нервного волокна по обеим сторонам от места раздражения находятся в состоянии покоя. В натуральных же условиях первый ПД, возникший на мембране тела нейрона, инициирует распространение возбуждения только в одном направлении — по аксону к другой клетке.
Сравнение механизма непрерывного и сальтаторного проведения возбуждения показывает, что различие между ними не принципиально. Оно заключается лишь в том, что очередные ПД в безмякотном волокне возникают на более близком расстоянии друг от друга, поскольку ионные каналы расположены в непосредственной близости друг от друга и непрерывно по всей длине нервного волокна. Поэтому такое проведение и назвали непрерывным.
Число одновременно возникающих ПД в мякотном волокне, в отличие от безмякотного, строго ограничено числом возбужденных перехватов Ранвье — максимально 5. В реальной же действительности ПД не «перепрыгивают» ни в мякотном, ни в безмякотном волокне — они возникают заново.
Однако сальтаторное проведение возбуждения имеет два важных преимущества перед непрерывным. Во-первых, сальтаторное проведение более экономично с точки зрения затрат энергии, так как возбуждаются только перехваты Ранвье, площадь которых менее 1% мембраны, и, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов Na+ и К+, расходующихся в процессе возникновения ПД (в миелинизированных волокнах при распространении возбуждения теряется ионов в 100 раз меньше, чем в немиелинизированных). Во-вторых, возбуждение в миелинизированных волокнах проводится с большей скоростью (см. таблицу), чем в безмиелиновых волокнах, так как в них электрическое поле ПД распространяется значительно дальше на соседние перехваты Ранвье, поскольку электроизоляция (миелиновые муфты) уменьшает рассеивание электрического поля.
В процессе проведения возбуждения время затрачивается только на перпендикулярное относительно мембраны волокна движение ионов в клетку и из клетки при формировании нового ПД, а влияние электрического поля возникших ПД на соседний участок распространяется вдоль длины волокна мгновенно — время распространения электрического поля практически равно нулю. Скорость распространения возбуждения увеличивается также при большой амплитуде ПД, что является следствием формирования более сильного электрического поля, обеспечивающего критический уровень деполяризации нервного волокна на большем расстоянии и быстрее.
В настоящее же время одни физиологи считают, что возникший ПД электротонически вызывает возбуждение в соседнем участке нервного или мышечного волокна; по мнению других, W. Willis (2004) и R. Klinke (2004), это осуществляется с помощью локальных входящих и выходящих токов ионов и между соседними участками волокна в продольном направлении. Однако электротон — это частичная деполяризация мембраны, еще не активирующая ионные каналы, поэтому в первом случае возбуждение вообще не возникло бы, а во втором случае оно распространялось бы слишком медленно.
Скорость распространения электрического поля хорошо иллюстрируется процессом общения по телефону — собеседники могут находиться на расстоянии нескольких тысяч километров друг от друга, но слова они слышат через такой же промежуток времени, как и при непосредственном разговоре.
Характеристика проведения возбуждения по нервному волокну. 1. Двустороннее проведение возбуждения можно продемонстрировать в эксперименте при нанесении раздражения в любом участке нерва или нервного волокна, при этом возбуждение регистрируется как в одном, так и в другом направлении от места раздражения.
2. Изолированное проведение возбуждения в отдельных волокнах нервного ствола обусловлено тем, что влияние электрического поля ПД соседнего волокна не возбуждает другие волокна нерва вследствие изолирующего эффекта их оболочек и интерстиция. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияния функционально разнородных волокон нерва. Однако если одновременно возбуждается значительное количество волокон, то возникает достаточно сильное электрическое поле, способное открыть ворота натриевых каналов соседних и таким образом усилить нервное влияние на эффекторные клетки или нейроны в регуляторных процессах.
3. Бездекрементное (без затухания; лат. decrementum — убывание, уменьшение) проведение по всей длине нервного волокна, так как ПД возникают в каждом участке волокна заново под влиянием предыдущего ПД, и его величина в каждом участке волокна складывается из величины ПП и фазы инверсии ПД.
4.Большая скорость проведения возбуждения (до 120 м/с) в нервных волокнах типа Аα. Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока — от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (время полного кругооборота крови — около 22 с). Большая скорость распространения ПД обеспечивает быстрое влияние на другие нейроны, рабочие органы, получение обратной информации, играющей важную роль в регуляторных процессах.
Малая утомляемость нервного волокна впервые была показана Н.Е. Введенским (1883): в проводимых опытах нерв сохранял способность к проведению возбуждения в течение 6—8 ч непрерывного раздражения несильными импульсами тока при условии наличия кислорода в окружающей среде и поддержания влажного состояния нерва. Это обусловлено тем, что при проведении ПД по нервным волокнам используется незначительная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для их восстановления. Расход энергии в нервном волокне на единицу массы в миллион раз меньше, чем в работающей мышце.
Высокая лабильность — нервное волокно может проводить до 500—1000 имп./с.
Возбуждение от нервного волокна передается к другой клетке с помощью синапса.
Дата добавления: 2016-02-16; просмотров: 5260;