Механизмы переноса электрона при фотосинтезе. Сопряжение окислительно-восстановительных реакций с трансмембранным переносом протона
Последовательность отдельных реакций в фотобиологических процессах включает следующие стадии: поглощение кванта света хромофорной группой и образование электронно-возбужденных состояний -> миграция энергии электронного возбуждения-> o первичный фотофизический акт и появление первичных фотопродуктов —> образование первичных стабильных химических соединений -> физиолого-биохимические процессы -> конечный фотобиологический эффект.
В основе первичных процессов фотосинтеза лежит сложная совокупность окислительно- восстановительных реакций переноса электрона в электрон-траспортной цепи (ЭТЦ). (Картинка Z-схема) 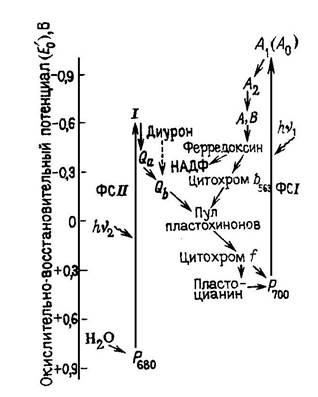
ФС2 и ФС1 функционируют последовательно: донором электронов для ФС1 служат восстановленные в результате действия ФС2 фотопродукты. Дальний красный свет (лямбда > 680нм) поглощается преимущественно пигментами ФС1 и вызывает окисление цитохрома, который восстанавливается ФС2 при поглощении коротковолнового света (дл.волны < 680нм). Оптимальная интенсивность фотосинтеза наблюдается при определенном соотношении между количеством возбужденных ФС1 и ФС2, которое зависит от спектрального состава света. Поглощение света происходит пигментами светособирающего (СС) пигмент - белкового (ПБ) комплекса (ССПБК), от которого, как из резервуара, энергия возбуждения передается на пигмент - белковые комплексы ФС1 и ФС2 (ПБК1 и ПБК2) и далее непосредственно к реакционным центрам РЦ1 и РЦ2. В зависимости от конформационного состояния фотосинтетических мембран изменяется топография расположения ПБК1 и ПБК2 , их связь с ССПБК, и распределение энергии возбуждения между ФС1 и ФС2. Это определяется присутствием ионов в среде, рН среды, степенью фосфорилирования и поверхностным зарядом белков ССПБК.
Основной принцип работы РЦ заключается в том, что при попадании возбуждения от (ПБК) в фотоактивный пигмент (Р) самого РЦ происходят отрыв электрона и восстановление первичного акцептора (А1), который затем передает электрон следующему акцептору (А2) и так далее по ЭТЦ. (картинка - передача е) 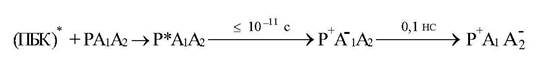
Восстановление Р+ происходит затем от доноров электрона в ЭТЦ. В качестве A1 в РЦ выступают производные порфириновых молекул (например, феофитин в РЦ2), а роль вторичного акцептора выполняют молекулы хинона (РЦ пурпурных бактерий), пластохинона (РС2) или железосерные белки (РЦ1). Эффективность отрыва электрона от Р+ и переноса его на А1 достигает 95 - 98% и происходит за времена несколько пикосекунд (1пс=10-12 с). Если в РЦ исходно в темноте вторичный акцептор химически восстановлен, то после появления электрона на А1 происходят его рекомбинация с Р+ за 5 - 10 нс и появление Р+. Это сопровождается замедленным свечением. (картинка – замедленное свечение). 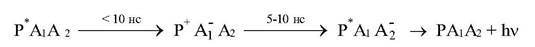
В РЦ пурпурных бактерий кинетика переноса электрона между Р (димер бактериохлорофилла), A1 (бактериофеофитин БФФ), А2 (первичный хинон QA), Аз (вторичный хинон Qb)
– 
Общая схема первичных процессов бактериального типа,
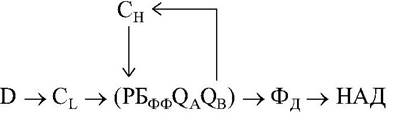
где внешним источником электронов для восстановления НАД служит сереводород, органические молекулы. Цитохром СН обеспечивает циклический поток электронов, а цитохром Cl - связь циклического переноса с внешними донорами электронов. + картинка реакции в РЦ пурпурных бактерий. 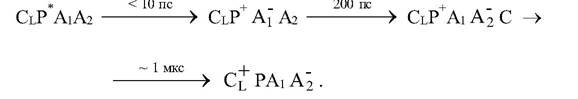
Важнейшая особенность функционирования РЦ состоит в том, что отдельные стадии переноса электрона могут протекать эффективно и при низких температурах, вплоть до температур жидкого азота и жидкого гелия.
Первичный хинон, получив в РЦ электрон, изменяет характер своего движения и переходит на другую конформационную координату, соответствующую его восстановленному состоянию. Здесь он, достигнув определенного контактного состояния со вторичным хиноном, отдает ему электрон. При низких температурах подвижность первичного хинона падает, а следовательно, уменьшается и эффективность переноса электрона от него на вторичный хинон. Если, однако, образец медленно охлаждать в условиях постоянного интенсивного освещения, когда в результате действия света и отрыва электрона от Р молекулы первичного хинона в основном находятся в восстановленном состоянии, то картина существенно изменяется.
Картинка «передача е на пул Пх». 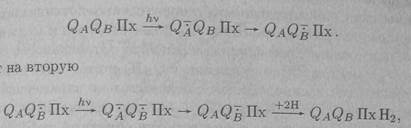 Семихиноныхарактеризуются поглощением в спектральной области 320-450 нм (первая вспышка). После второй вспышки света происходит образование дважды восстановленного хинона Q2-B и исчезновение полосы поглощения семихинона. Молекула Q2-B протонируется из внешнего окружения и обменивается с одной молекулой хинона из пула Пх, занимая ее место. Система возвращается в исходное состояние с той лишь разницей, что одна молекула пула теперь протонирована: ПхН2. Третья вспышка опять приводит к образованию семихинонных радикалов и соответствующему увеличению поглощая. Картинка «общая схема процесса».
Семихиноныхарактеризуются поглощением в спектральной области 320-450 нм (первая вспышка). После второй вспышки света происходит образование дважды восстановленного хинона Q2-B и исчезновение полосы поглощения семихинона. Молекула Q2-B протонируется из внешнего окружения и обменивается с одной молекулой хинона из пула Пх, занимая ее место. Система возвращается в исходное состояние с той лишь разницей, что одна молекула пула теперь протонирована: ПхН2. Третья вспышка опять приводит к образованию семихинонных радикалов и соответствующему увеличению поглощая. Картинка «общая схема процесса». 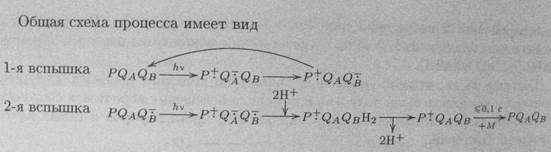 Образованный дважды восстановленный вторичный хинон QвH2 быстро окисляется при достаточно высоких окислительно-восстановительных потенциалах среды. Семихинонный вторичный акцептор более стабилен и медленно окисляется за счет реакции с окисленной формой медиатора.
Образованный дважды восстановленный вторичный хинон QвH2 быстро окисляется при достаточно высоких окислительно-восстановительных потенциалах среды. Семихинонный вторичный акцептор более стабилен и медленно окисляется за счет реакции с окисленной формой медиатора.
| <== предыдущая лекция | | | следующая лекция ==> |
| | | Кинетика изменений потоков ионов при потенциале действия. Описание ионных потоков в модели Ходжкина-Хаксли |
Дата добавления: 2016-04-11; просмотров: 2102;