Кинетика изменений потоков ионов при потенциале действия. Описание ионных потоков в модели Ходжкина-Хаксли
Тут в разных источниках все разными буквами обозначают. Я придерживался всю дорогу одной системы, ноиногда вместо φ где-то пишут Е... А где-то нет=)
Тут можно по-всякому подступиться. Я все-таки не буду тут описывать выведение уравнение Гольдмана-Ходжкина-Хаксли, потому что вопрос тут более философский. Если что, см. 3.1.
Для начала, надо написать, чему равны равновесные потенциалы по данному иону. Для этого надо рассмотреть вечное корыто с полупроницаемой мембраной, в одну половинку налили электролит. И написать, чему равен электрохимический потенциал в каждой половине для какого-нибудь иона (это все подробно есть в 3.1.).
µin= µ0+RTlnCin+zFφin и µout= µ0+RTlnCout+zFφout. У нас равновесие, так что эти потенциалы равны. Запишем это, приравняем и получим уравнение Нернста для одного иона – это и есть равновесный потенциал по данному иону: φi=RT/(zF)* ln(Cin/Cout) (тут и далее под φ надо понимать разность потенциалов снаружи минус внутри. Дело в том, что правильнее говорить равновесная разность потенциалов, но поскольку привыкли говорить просто равновесный потенциал, то и вместо Δφ пишут φ, хотя по факут это неправильно).
Уравнение Гольдмана-Ходжкина-Хаксли описывает, чему равен потенциал на мембране (для нервных волокон и мышц)
φ=RT/F*ln((PK[Kout]+ PNa[Naout]+ PCl[Clin])/( PK[Kin]+ PNa[Nain]+ PCl[Clout])).
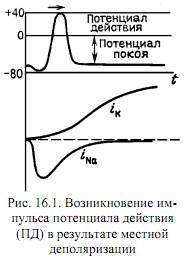
Во время покоя все проницаемости постоянны, поэтому ПП не меняется. Однако при возбуждении мембраны (при электростимуляции или при воздействии медиатором) открываются Na-каналы, и PNa бешено растет. Числитель увеличивается, φ растет, и мы наблюдаем фазу деполяризации, то есть увеличение φ от отрицательного значания к нулю. Вслед за этим φ растет еще больше, уходя в положительные значения – овершут. Чтобы доказать, что дело тут в проницаемостях, напишу, что при ПП PK:PNa=1:0.04, а в момент, соответсвующий овершуту PK:PNa=1:20. (При ПП PNa<< PK и PCl, так что ПП это приблизительно среднее арифметическое между φK и φCl ( (-79-54)/2 ) – роль K и Cl в ПП.)
Итак, фаза деполяризации обусловлена током Na+ внутрь – роль Na в ПД! Во время этой фазы растет PNa и φ->φNa. Этот факт так же недурно будет проиллюстрировать другим взглядом на мембрану. Рассмотрим токи. Как известно, Ii=gi*(φ-φi). Если это расписать для всех трех ионов, а так же написать, что сумма всех токов в ПП равна нулю, то получится системы из 4х уравнений. Из нее легко вывести следующее: φ=(gKφK+gNaφNa+gClφCl)/(gK+gNa+ gCl). Тут очень хорошо видно, что с ростом проводимости g растет вклад конкретного равновесного потенциала в общий мембранный потенциал.
Однако, φ не растет бесконечно, по двум причинам. 1. Инактивация натриевых каналов (сразу после открытия они надолго закрываются). 2. Открываются K-каналы, и К идет внутрб. И наступает фаза РЕполяризации – роль K в ПД. Посколку Na-каналы инактивированы, то φ->φK. И приближается к нему даже больше, чем при ПП, т.к. вклад натрия меньше из-за инактивации натриевых каналов. Так что мы наблюдаем следовую гиперполяризацию (тожероль К в ПД).
Теперь про ПД в поперечно-полосатой мышце. Медиатор попал на мышцу в области под названием концевая пластинка (КП, она под синапсом). Связался с рецептором на канале (обычно это Н-холиновый рецептор), канал открылся, в саркоплазму потек Na+ (канал неселктивен, просто калию там делать нечего, его и так там дофига). Это вызовет генерацию ПКП – потенциала концевой пластинки (это не ПД, он не подчиняется приципу все или ничего). Он вокруг КП есть нормальные натриевые каналы, и если ПКП надпороговый, то за счет НИХ и сгенерируется ПД. Он дойдет до Т-трубочки, в ней откроется дигидроперидиновые потенциал зависимые кальциевые каналы, и уже Са пойдет внутрь. Он свяжется с рианодиновыми кальций зависимыми кальцевыми каналами на СПР, они откроются, и уже МНОГО Са пойдет в саркоплазму. Это вызовет сокращение (это уж не буду описывать...). Это – роль Ca в ПД.
Теперь про водоросль. Про хару, в частности. У нее 2 отличия: 1. ПП обеспечивается φK и работой Н-АТФаз. 2. Деполяризация – сначала током Са внутрь, а в ответ на эту деполяризацию дело еще усугубляется открытыем хлорных каналов, и хлор течет наружу, еще больше деполяризуя мембрану. А натрий тут воооообще ничего не решает.
Теперь про токи. В данном вопросе под потоками имеются в виду, конечно же, токи=) Во-первых, они описываются этим вот уравнением: Ii=gi*(φ-φi). То есть ток зависит от двух изменяющихся величин: потенциала на мембране и проводимости. Проводимость изменяется в зависимости от потенциала на мембране. Короче: Ii=f(φ, gi(φ)). φi – в данном случае константа. Известно, что gNa сначала быстро растет, а затем быстро экспоненциально убывает, а gK растет по S-образной кривой.
Вопрос о том, как gi управляется электрическим полем, вызванным потенцалом на мембране, - ключевой в биофизике мембранных процессов. В модели Х-Х предполагается, что проводимость регулируется некоторыми положительно заряженными частицами, которые перемещаются в мембране при изменении поля. Эти частицы соответсвующим образом закрывают или открывают ионным канал. Для К-каналов считается, что есть 4 активирующие частицы, а для Na-каналов 3 активирующие и 1 инактивирующая. Это возволило создать математический аппарат, с высокой точностью моделирующий нервный импульс. В функционально структуре канала удалось выделить элементы, отвечающие за механизмы селекции ионов – селективный фильтр, - активации – активационные ворота – и инактивации – инактивационные ворота. Эти управляющие частицы называются воротными частицами. Их движение можно обнаружить эксперементально по возникновению т.н. воротных токов. Они появляются в результате смещения частиц в мембране под влиянием наложенного на мембрану электрического потенциала. Удалось обнаружить воротные токи смещения, связанные с частицами, открывающими Na-канал. Предполагается, что эти токи вызваны изменением ориентации диполей, выстилающих внутреннюю полость канала.
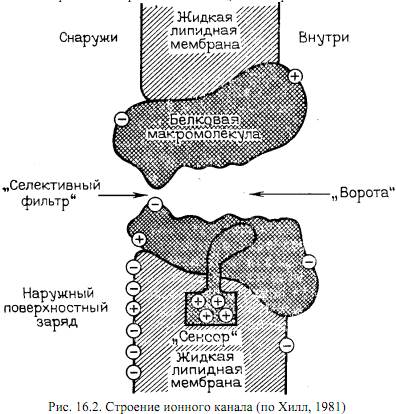
Полярные группы, определяющие диполыюе окружение иона в канале, влияют на энергию иона и на прохождение его через канал, Если иод влиянием деполяризующего электрического импульса изменяется их ориентация, то это вызовет смещение заряженных групп и изменение проводимости канала. Процесс переориентации диполей может носить кооперативный характер и быть достаточно резким. В этом случае энергия, необходимая для переориентации каждого элементарного диполя, должна зависеть не только от его собственной энергии, но и от доли диполей, уже изменивших свою ориентацию. Иными словами, по мере изменения ориентации части диполей энергия, необходимая для переориентации оставшихся диполей, уменьшается тем значительнее, чем больше число уже переориентированных диполей. Можно представить себе, что в исходном состоянии диполи мешают друг другу изменить ориентацию под действием поля, а переориентированные диполи уже "не путаются иод ногами" у оставшихся. В результате такого рода кооперативного эффекта проводимость канала очень быстро "лавинообразно" нарастает под действием приложенного электрического импульса. Заметим, что и конформацнонные перестройки в канале, сопровождающиеся поворотом диполей, могут также приводить к скачкообразным изменениям проводимости в одиночном канале. Фактически во всех предложенных моделях речь идет о своего рода фазовых переходах в мембранах, лежащих в основе скачкообразных переходов канала между двумя состояниями.
Дата добавления: 2016-04-11; просмотров: 1947;