Соотношение между нагрузкой и скоростью укорочения мышцы.
Основные характеристики мышечной деятельности
Скелетные мышцы.
Скелетные (поперечнополосатые) мышцы — это "машины", преобразующие химическую энергию непосредственно в механическую и тепловую. Сокращение мышц возникает в ответ на электрические импульсы, приходящие к ним от а- мотонейронов — нервных клеток, лежащих в передних рогах спинного мозга. Мышцы и иннер-вирующие их мотонейроны составляют нервно-мышечный аппарат человека. В результате сократительной деятельности скелетных мышц осуществляется поддержание позы человека, перемещение частей тела относительно друг друга, передвижение человека в пространстве.
Основным морфо-функциональным элементом нервно-мышечного аппарата является двигательная единица (ДЕ). ДЕ — это мотонейрон с иннервируемыми им мышечными волокнами. Аксон мотонейрона из спинного мозга проходит в составе периферических нервов до мышцы, внутри которой разветвляется на множество концевых веточек. Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно-мышечный синапс. Импульсы, идущие по аксону мотонейрона, активируют все иннервируемые им мышечные волокна. Поэтому ДЕ функционирует как единое морфофунк-циональное образование.
Скелетная мышца состоит из пучков вытянутых в длину клеток — мышечных волокон, обладающих тремя свойствами: возбудимостью, проводимостью и сократимостью (см.главу I). Отличительной чертой мышечных клеток от клеток, не обладающих свойством сократимости, является наличие саркоплазматического ретикулума. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. В мембране саркоплазматического ретикулума находятся две транспортные системы, обеспечивающие освобождение от ретикулума ионов кальция при возбуждении и их возврат из миоплазмы обратно в ретикулум при расслаблении мышцы. В механизме освобождения ионов кальция из ретикулума при возбуждении мышечной клетки важную роль играет система поперечных трубочек (Т-система), представляющих собой впячивания поверхностной мембраны мышечного волокна. К противоположным сторонам поперечной трубочки примыкают боковые цистерны ретикулума. Две терминальные цистерны ретикулума вместе с трубочкой образуют так называемую триаду — анатомическую
146
структуру, в зоне которой нервные импульсы, распространяющиеся по поперечным трубочкам вглубь мышечного волокна, запускают процесс выхода ионов кальция из саркоплазматического ретикулума и, следовательно, всю последующую цепочку изменений, приводящую, в конечном итоге, к развитию сокращения мышцы.
Мышечные волокна имеют диаметр от 10 до 100 мкм и длину от 5 до 400 мм (в зависимости от длины мышцы). В каждом мышечном волокне содержится до 1000 и более сократительных элементов ми-офибрилл, толщиной 1-3 мкм. Каждая миофибрилла состоит из множества параллельно лежащих толстых и тонких нитей — миофиламен-тов. Толстые нити состоят из молекул белка миозина, а тонкие — из белка актина. Миозиновые нити имеют отходящие от них биполярно поперечные выступы около 20 нм, с головками, состоящими примерно из 150 молекул миозина. Во время сокращения каждая головка миозина, или поперечный мостик, может связывать миозиновую нить с соседней актиновой. Кроме того, в состав тонких нитей входят еще два белка — тропонин и тропомиозин, необходимые для развития процессов сокращения и расслабления мышцы.
Расположение миозиновых и тонких актиновых белковых нитей строго упорядочено (рис.4.1.). Пучок лежаших в середине саркомера нитей миозина выглядит в световом микроскопе как темная полоска. Благодаря свойству двойного лучепреломления в поляризованном свете (то есть анизотропии) она называется А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и поэтому выглядят светлыми. Эти изотропные J-диски тянутся до Z-пластин. Благодаря такому периодическому чередованию светлых и темных полос миофибриллы скелетной мышцы
 | |||
 | |||
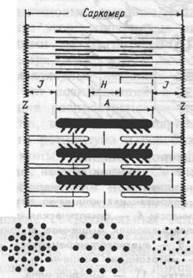 Рис.4.1. Схема саркомера мышечного волокна и взаимного расположения толстых миозиновых и тонких актиновых миофиламентов.
Рис.4.1. Схема саркомера мышечного волокна и взаимного расположения толстых миозиновых и тонких актиновых миофиламентов.
Поперечный срез миофибриллы дает
представление о гексагональном
распределении актиновых и миозиновых
нитей.
Z — линии, разделяющие два соседних
саркомера;
J — изотропный диск;
А — анизотропный диск;
Н — участок с уменьшенной
анизотропностью.






 выглядят исчерченными (поперечно-полосатыми). Если мышца расслаблена, то в средней части А-диска различается менее плотная Н- зона, состоящая только из толстых миофиламентов. Н- зона не просматривается во время сокращения мышцы. По середине J-дис-ка проходит темная полоска — это Z линия. Участок миофибриллы между двумя Z линиями называется саркомером.
выглядят исчерченными (поперечно-полосатыми). Если мышца расслаблена, то в средней части А-диска различается менее плотная Н- зона, состоящая только из толстых миофиламентов. Н- зона не просматривается во время сокращения мышцы. По середине J-дис-ка проходит темная полоска — это Z линия. Участок миофибриллы между двумя Z линиями называется саркомером.
Передача возбуждения в нервно-мышечном синапсе.Структура нервно- мышечного синапса представлена на рисунке 4.2. В процессе передачи возбуждения с нерва на мышечные волокна выделяют три последовательных процесса: 1. электрический, включащий достижение нервным импульсом концевой веточки аксона, деполяризацию и повышение проницаемости ее мембраны, выделение аце-тилхолина (АХ) в синаптическую щель; 2. химический, основу которого составляет диффузия медиатора АХ к постсинаптической мембране и образование на ней его комплекса с холинорецептором; 3. электрический, включащий увеличение ионной проницаемости постсинаптической мембраны, возникновение локального электри-
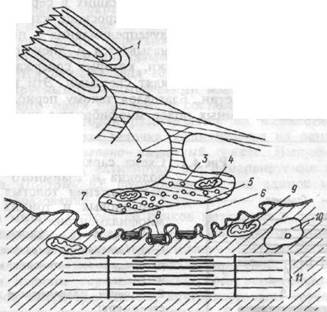
Рис.4.2. Схема элементов нервно- мышечного синапса.
1 — миэлиновая оболочка аксона; 2 — концевые веточки аксона; 3 — пузырьки, содержащие ацетилхолин; 4 — митохондрия; 5 — пресинаптическая мембрана, покрывающая концевую веточку аксона в зоне нервно-мышечного синапса; 6 — синаптическая щель; 7 —постсинаптическая мембрана, покрывающая мышечное волокно в зоне нервно-мышечного синапса; 8 — ацетилхолинорецепторы на постсинаптической мембране; 9 — митохондрия мышечного волокна; 10 — ядро мышечной клетки; 11 — миофибрилла.
 148
148

 ческого потенциала (потенциала концевой пластинки; ПКП), развитие потенциала действия мышечного волокна.
ческого потенциала (потенциала концевой пластинки; ПКП), развитие потенциала действия мышечного волокна.
Запасов АХ в нервном окончании достаточно для проведения лишь примерно 10 000 импульсов. При длительной же импульсации мотонейрона, несмотря на постоянный синтез АХ (см. главу 3), его содержание в концевых веточках может постепенно уменьшаться. В результате этого возможны нарушения передачи возбуждения в нервно-мышечных синапсах — пресинаптический нервно-мышечный блок.
Временно возникающий на постсинаптической мембране комплекс "АХ- рецептор" после прохождения каждого импульса разрушается ферментом ацетил-холинэстеразой. Однако при длительной высокочастотной импульсации мотонейрона (например при длительной и напряженной мышечной работе) АХ не успевает разрушаться и накапливается в синаптической щели. Способность постсинаптической мембраны к генерации ПКП при этом снижается и развивается частичный или полный постсинаптический нервно- мышечный блок, приводящий либо к частичному, либо даже полному прекращению развития потенциалов действия на мембране мышечного волокна.
Механизмы сокращения мышечного волокна.В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым ми-офиламентам. Тропомиозин расположен таким образом, что блокирует участки актина, способные взаимодействовать с поперечными мостиками миозина. Тропонин тормозит миозин — АТФ-азную активность и поэтому АТФ не расщепляется. Мышечные волокна находятся в расслабленном состоянии.
При сокращении мышцы длина А-дисков не меняется, J-диски укорачиваются, а Н-зона А-дисков может исчезать (рис. 4.3.). Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения (теорией скольжения) тонких актиновых миофиламентов вдоль толстых миозиновых. В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми. Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазмати-ческого ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения. Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют "электромеханическим сопряжением". Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке 4.4.
149





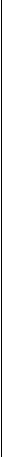


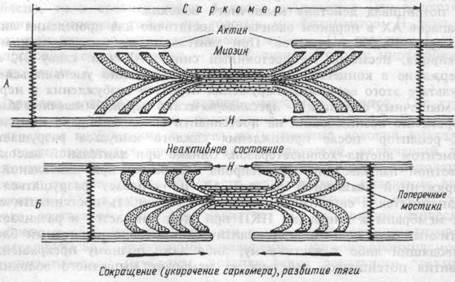
Рис.4.3. Сокращение мышцы.
А. Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии.
Б. Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
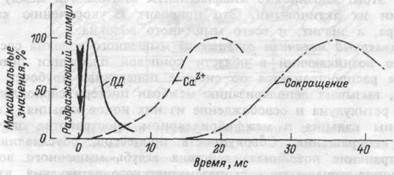
Рис.4.4. Схема временной последовательности развития
потенциала действия (ПД), освобождения ионов кальция (Са2+) и развития изометрического сокращения мышцы.
 150
150

 При концентрации ионов Са2+ в межмиофибриллярном пространстве ниже 10" тропомиозин располагается таким образом, что блокирует прикрепление поперечных миозиновых мостиков к нитям актина. Поперечные мостики миозина не взаимодействуют с нитями актина. Продвижение относительно друг друга нитей актина и миозина отсутствует. Поэтому мышечное волокно находится в расслабленном состоянии. При возбуждении волокна Са2+ выходит из цистерн саркоплаз-матического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са2+ молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают "гребко-вые" движения в сторону центра саркомера происходит "втягивание" актиновых миофиламентов в промежутки между толстыми миозиновы-ми нитями и укорочение мышцы.
При концентрации ионов Са2+ в межмиофибриллярном пространстве ниже 10" тропомиозин располагается таким образом, что блокирует прикрепление поперечных миозиновых мостиков к нитям актина. Поперечные мостики миозина не взаимодействуют с нитями актина. Продвижение относительно друг друга нитей актина и миозина отсутствует. Поэтому мышечное волокно находится в расслабленном состоянии. При возбуждении волокна Са2+ выходит из цистерн саркоплаз-матического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са2+ молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают "гребко-вые" движения в сторону центра саркомера происходит "втягивание" актиновых миофиламентов в промежутки между толстыми миозиновы-ми нитями и укорочение мышцы.
Источником энергии для сокращения мышечных волокон служит АТФ. С инактивацией тропонина ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что обеспечивает энергией поперечные мостики. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. На миозиновом поперечном мостике образуется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снижается до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
При однократном движении поперечных мостиков вдоль актиновых нитей (гребковых движениях) саркомер укорачивается примерно на 1% его длины. Следовательно, для полного изотонического сокращения мышцы необходимо совершить около 50 таких гребковых движений. Только ритмическое прикрепление и отсоединение головок миозина может втянуть нити актина вдоль миозиновых и совершить требуемое укорочение целой мышцы. Напряжение, развиваемое мышечным волокном, зависит от числа одновременно замкнутых поперечных мостиков. Скорость развития напряжения или укорочения волокна определяется частотой замыкания поперечных мостиков, образуемых в единицу времени, то есть скоростью их прикрепления к актиновым миофиламентам. С увеличением скорости укорочения мышцы число одновременно прикрепленных поперечных мостиков в каждый момент времени уменьшается. Этим и можно объяснить уменьшение силы сокращения мышцы с увеличением скорости ее укорочения.
При одиночном сокращении процесс укорочения мышечного волокна заканчивается через 15-50 мс, так как активирующие его ионы кальция возвращаются при помощи кальциевого насоса в
151

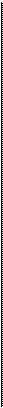

 цистерны саркоплазматического ретикулума. Происходит расслабление мышцы.
цистерны саркоплазматического ретикулума. Происходит расслабление мышцы.
Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии. Ее источником служит АТФ. Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны. При снижении содержания ионов кальция до подпорогового уровня (ниже 10 V) молекулы тропонина принимают форму, характерную для состояния покоя. При этом вновь тропомиозин блокирует участки для прикрепления поперечных мостиков к нитям актина. Все это приводит к расслаблению мышцы вплоть до момента прихода очередного потока нервных импульсов, когда описанный выше процесс повторяется. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.
Режимы и типы мышечных сокращений.Режим сокращений мышечных волокон определяется частотой импульсации мотонейронов. Механический ответ мышечного волокна или отдельной мышцы на однократное их раздражение называется одиночным сокращением. При одиночном сокращении выделяют: 1. фазу развития напряжения или укорочения; 2. фазу расслабления или удлинения (рис.4.5.). Фаза расслабления продолжается примерно в два раза дольше, чем фаза напряжения. Длительность этих фаз зависит от морфофункциональ-ных свойств мышечного волокна: у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения составляет 7-10 мс, а у наиболее медленных волокон камбаловидной мышцы — 50-100 мс.
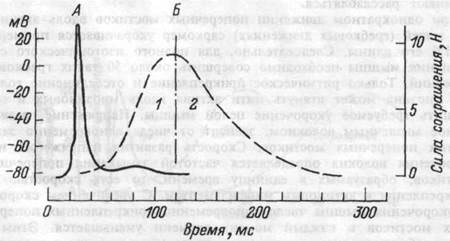
Рис.4.5. Развитие во времени потенциала действия (А) и изометрического сокращения мышцы, приводящей большой палец кисти (Б).
1 — фаза развития напряжения; 2 — фаза расслабления.
 152
152




 В естественных условиях мышечные волокна двигательной единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон. Так, режим одиночного сокращения медленных волокон камбаловидной мышцы человека обеспечивается при частоте импульсации мотонейрона менее 10 имп/с, а быстрых волокон глазодвигательных мышц — при частоте импульсации мотонейрона менее 50 имп/с.
В естественных условиях мышечные волокна двигательной единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон. Так, режим одиночного сокращения медленных волокон камбаловидной мышцы человека обеспечивается при частоте импульсации мотонейрона менее 10 имп/с, а быстрых волокон глазодвигательных мышц — при частоте импульсации мотонейрона менее 50 имп/с.
В режиме одиночного сокращения мышца способна работать длительное время без развития утомления. Однако в связи с тем, что длительность одиночного сокращения невелика, развиваемое мышечными волокнами напряжение не достигает максимально возможных величин. При относительно высокой частоте импульсации мотонейронов каждый последующий раздражающий импульс приходится на фазу предшествующего напряжения волокона, то есть до того момента, когда оно начинает расслабляться. В этом случае механические эффекты каждого предыдущего сокращения суммируются с последующим. Причем величина механического ответа на каждый последующий импульс меньше, чем на предыдущий. После нескольких первых импульсов последующие ответы мышечных волокон не изменяют достигнутого напряжения, а лишь поддерживают его. Такой режим сокращения называется гладким тетанусом (рис.4.6.). В подобном режиме двигательные единицы мышц человека работают при развитии максимальных изометрических усилий. При гладком тетанусе развиваемое ДЕ напряжение в 2-4 раза больше, чем при одиночных сокращениях.
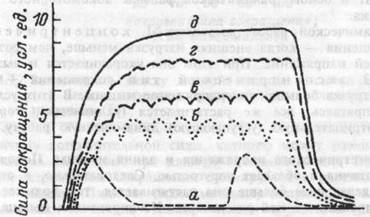
Рис.4.6. Одиночные (а) и тетанические (б,в,г,д) сокращения скелетной мышцы. Накладывание волн сокращения друг на друга и образование тетануса при частотах раздражения: 5—15 раз в 1 с; в — 20 раз в 1 с; г — 25 раз в 1 с; д — более 40 раз в 1 с (гладкий тетанус).
 153
153




 В тех случаях, когда промежутки между последовательными импульсами мотонейрона меньше времени полного цикла одиночного сокращения, но больше длительности фазы напряжения, сила сокращения ДЕ колеблется. Этот режим сокращения называется зубчатым тетанусом (рис. 4.6.).
В тех случаях, когда промежутки между последовательными импульсами мотонейрона меньше времени полного цикла одиночного сокращения, но больше длительности фазы напряжения, сила сокращения ДЕ колеблется. Этот режим сокращения называется зубчатым тетанусом (рис. 4.6.).
Гладкий тетанус для быстрых и медленных мыши достигается при разных частотах импульсации мотонейронов. Зависит это от времени одиночного сокращения. Так, гладкий тетанус для быстрой глазодвигательной мышцы проявляется при частотах свыше 150-200 имп/с, а у медленной камбаловидной мышцы — при частоте около 30 имп/с. В режиме тетанического сокращения мышца способна работать лишь короткое время. Это объясняется тем, что из-за отсутствия периода расслабления она не может восстановить свой энергетический потенциал и работает как бы "в долг".
Механическая реакция целой мышцы при ее возбуждении выражается в двух формах — в развитии напряжения и в укорочении. В естественных условиях деятельности в организме человека степень укорочения мышцы может быть различной. По величине укорочения различают три типа мышечного сокращения: 1. изотонический — это сокращение мышцы, при которой ее волокна укорачиваются при постоянной внешней нагрузке. В реальных движениях чисто изотоническое сокращение практически отсутствует; 2. изометрический — это тип активации мышцы, при котором она развивает напряжение без изменения своей длины. Изометрическое сокращение лежит в основе статической работы; 3. ауксотони-ч е с к и й или анизотонический тип — это режим, в котором мышца развивает напряжение и укорачивается. Именно такие сокращения имеют место в организме при естественных локомоциях — ходьбе, беге и т.д. Изотонический и анизотонический типы сокращения лежат в основе динамической работы локомоторного аппарата человека.
При динамической работе выделяют: 1. концентрический тип сокращения — когда внешняя нагрузка меньше, чем развиваемое мышцей напряжение. При этом она укорачивается и вызывает движение; 2. эксцентрический тип сокращения — когда внешняя нагрузка больше, чем напряжение мышцы. В этих условиях мышца, напрягаясь, все же растягивается (удлиняется), совершая при этом отрицательную (уступающую) динамическую работу.
Сила изометрического напряжения и длина мышцы.Покоящаяся мышца эластична и обладает упругостью. Следовательно, в определенных пределах, чем больше она растягивается, тем большее продольное напряжение в ней развивается. Изолированная мышца имеет равновесную длину, при которой ее упругое напряжение равно нулю. Зависимость между длиной мышцы и ее напряжением в покое называется кривой пассивного напряжения (рис.4.7.). Кривая напряжения нарастает тем круче, чем больше степень растяжения мышцы.
Степень предварительного растяжения определяет не только величину пассивного эластического напряжения покоящейся мышцы, но
154



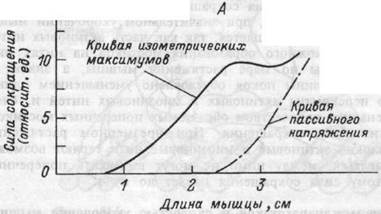
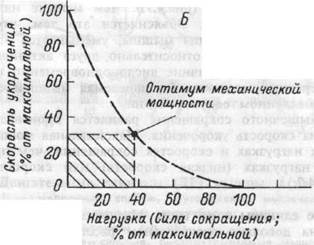
Рис.4.7. Зависимость между длиной мышцы и силой сокращения (А); между скоростью укорочения и величиной внешней нагрузки (Б).
 и величину дополнительной силы, которую может развивать мышца в случае ее активации при данной исходной длине. Прирост силы при изометрическом сокращении суммируется с пассивным напряжением мышцы. Пиковые (максимальные) напряжения в этих условиях называют максимум изометрического напряжения (рис.4.7.). Напряжение сокращающейся мышцы максимально, если ее длина составляет примерно 120 % от равновесной. Это состояние носит название длины покоя.
и величину дополнительной силы, которую может развивать мышца в случае ее активации при данной исходной длине. Прирост силы при изометрическом сокращении суммируется с пассивным напряжением мышцы. Пиковые (максимальные) напряжения в этих условиях называют максимум изометрического напряжения (рис.4.7.). Напряжение сокращающейся мышцы максимально, если ее длина составляет примерно 120 % от равновесной. Это состояние носит название длины покоя.
Укорочение мышцы меньше длины покоя или ее растяжение больше этой величины приводит к снижению силы сокращения. Причина этого состоит в специфике взаимодействия актиновых и
155




 миозиновых нитей. Поскольку напряжение, которое развивают ми-офибриллы в процессе развития сокращения, зависит от числа поперечных замкнутых мостиков, при значительном укорочении мышцы сила ее сокращения уменьшается, так как часть актиновых нитей выходит из зоны возможного образования мостиков на нитях миозина. Снижение силы по мере растяжения мышцы, а значит и саркомеров, больше длины покоя обусловлено уменьшением длины зон взаимного перекрытия актиновых и миозиновых нитей и, следовательно, меньшим количеством образуемых поперечных мостиков, обеспечивающих силу сокращения. При чрезмерном растяжении мышечного волокна актиновые и миозиновые нити теряют возможность перекрываться, между ними не могут возникать поперечные мостики. Поэтому сила сокращения падает до нуля.
миозиновых нитей. Поскольку напряжение, которое развивают ми-офибриллы в процессе развития сокращения, зависит от числа поперечных замкнутых мостиков, при значительном укорочении мышцы сила ее сокращения уменьшается, так как часть актиновых нитей выходит из зоны возможного образования мостиков на нитях миозина. Снижение силы по мере растяжения мышцы, а значит и саркомеров, больше длины покоя обусловлено уменьшением длины зон взаимного перекрытия актиновых и миозиновых нитей и, следовательно, меньшим количеством образуемых поперечных мостиков, обеспечивающих силу сокращения. При чрезмерном растяжении мышечного волокна актиновые и миозиновые нити теряют возможность перекрываться, между ними не могут возникать поперечные мостики. Поэтому сила сокращения падает до нуля.
Соотношение между нагрузкой и скоростью укорочения мышцы.
Скорость сокращения мышцы (то есть величина ее укорочения в единицу времени) зависит от величины внешней нагрузки, которую она вынуждена преодолевать (рис.4.7.). Чем меньше нагрузка, тем больше скорость сокращения. Объясняется это тем, что по мере увеличения скорости укорочения мышцы, уменьшается время взаимодействия скользящих друг относительно друга актиновых и миозиновых нитей. По этой причине число одновременно взаимодействующих поперечных мостиков и развиваемая мышцей сила меньше, чем при медленном ее укорочении.
Мощность мышечного сокращения равняется произведению мышечной силы на скорость укорочения. Максимальная мощность выше при умеренных нагрузках и скоростях сокращения, чем при близких к максимуму нагрузках (низкая скорость) или скоростях (малые нагрузки; рис.4.7.).
Двигательные единицы и их типы.Мышечные волокна каждой ДЕ расположены на довольно значительном расстоянии друг от друга. Число мышечных волокон, входящих в одну ДЕ, отличается в разных мышцах. Оно меньше в мелких мышцах, осуществляющих тонкую и плавную регуляцию двигательной функции (например, мышцы кисти, глаза) и больше вкрупных, не требующих столь точного контроля (икроножная мышца, мышцы спины). Так, в частности, в глазных мышцах одна ДЕ содержит 13-20 мышечных волокон, а ДЕ внутренней головки икроножной мышцы — 1500-2500. Мышечные волокна одной ДЕ имеют одинаковые морфофункциональные свойства.
По морфофункциональным свойствам ДЕ делятся на три основных типа (рис.4.8.): I — медленные, неутомляемые; ПА — быстрые, устойчивые к утомлению: II В — быстрые, легко утомляемые. Скелетные мышцы человека состоят из ДЕ всех трех типов. Одни из них включают преимущественно медленные ДЕ, другие — быстрые, третьи — и те, и другие.
Медленные, неутомляемые двигательные единицы (тип I). По сравнению с другими типами ДЕ у этих ДЕ наименьшие величины мотонейронов и, соответственно, наиболее низкие пороги их акти-
156






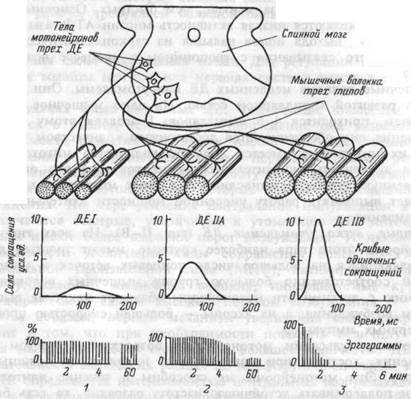
Рис.4.8. Двигательные единицы (ДЕ) мышцы и их типы.
1 — медленные, слабые, неутомляемые мышечные волокна.
Низкий порог активации мотонейрона;
2 — промежуточный тип ДЕ;
3 — быстрые, сильные, быстроутомляемые мышечные
волокна. Высокий порог активаций мотонейрона.
 вации, меньшие толщина аксона и скорость проведения возбуждения по нему. Аксон разветвляется на небольшое число концевых веточек и иннервирует небольшую группу мышечных волокон. У мотонейронов медленных ДЕ сравнительно низкая частота разрядов (6-10 имп/с). Они начинают функционировать уже при малых мышечных усилиях. Так, мотонейроны ДЕ камбаловидной мышцы человека при удобном стоянии работают с частотой 4 имп/с. Устойчивая частота их импульсации составляет 6- 8 имп/с. С повышением силы сокращения мышцы частота разрядов мотонейронов медленных ДЕ повышается незначительно. Мотонейроны медленных ДЕ способны поддерживать постоянную частоту разрядов в течение десятков минут.
вации, меньшие толщина аксона и скорость проведения возбуждения по нему. Аксон разветвляется на небольшое число концевых веточек и иннервирует небольшую группу мышечных волокон. У мотонейронов медленных ДЕ сравнительно низкая частота разрядов (6-10 имп/с). Они начинают функционировать уже при малых мышечных усилиях. Так, мотонейроны ДЕ камбаловидной мышцы человека при удобном стоянии работают с частотой 4 имп/с. Устойчивая частота их импульсации составляет 6- 8 имп/с. С повышением силы сокращения мышцы частота разрядов мотонейронов медленных ДЕ повышается незначительно. Мотонейроны медленных ДЕ способны поддерживать постоянную частоту разрядов в течение десятков минут.
Мышечные волокна медленных ДЕ развивают небольшую силу при сокращении в связи с наличием в них меньшего, по сравнению с
157

 быстрыми волокнами, количества миофибрилл. Скорость сокращения этих волокон в 1,5-2 раза меньше, чем быстрых. Основными причинами этого являются низкая активность миозин АТФ-азы и меньшие скорость выхода ионов кальция из саркоплазматического ре-тикулума и его связывания с тропонином в процессе возбуждения волокна.
быстрыми волокнами, количества миофибрилл. Скорость сокращения этих волокон в 1,5-2 раза меньше, чем быстрых. Основными причинами этого являются низкая активность миозин АТФ-азы и меньшие скорость выхода ионов кальция из саркоплазматического ре-тикулума и его связывания с тропонином в процессе возбуждения волокна.
Мышечные волокна медленных ДЕ малоутомляемы. Они обладают хорошо развитой капиллярной сетью. На одно мышечное волокно, в среднем, приходится 4- 6 капилляров. Благодаря этому во время сокращения они обеспечиваются достаточным количеством кислорода. В их цитоплазме имеется большое количество митохондрий и высокая активность окислительных ферментов. Все это определяет существенную аэробную выносливость данных мышечных волокон и позволяет выполнять работу умеренной мощности длительное время без утомления.
Быстрые, легко утомляемые ДЕ (тип П-В). Из всех типов ДЕ мотонейроны этого типа наиболее крупные, имеют толстый аксон, разветвляющийся на большое число концевых веточек и иннерви-рующий соответственно большую группу мышечных волокон. По сравнению с другими эти мотонейроны обладают наиболее высоким порогом возбуждения, а их аксоны — большей скоростью проведения нервных импульсов.
Частота импульсации мотонейронов возрастает с ростом силы сокращения, достигая при максимальных напряжениях мышцы 25-50 имп/с. Эти мотонейроны не способны в течение длительного времени поддерживать устойчивую частоту разрядов, то есть быстро утомляются.
Мышечные волокна быстрых ДЕ, в отличие от медленных, содержат большее число сократительных элементов — миофибрилл, поэтому при сокращении развивают большую силу. Благодаря высокой активности миозиновой АТФ-азы у них выше скорость сокращения. Волокна этого типа содержат больше гликолитических ферментов, меньше митохондрий и миоглобина, окружены меньшим, по сравнению с медленными ДЕ, количеством капилляров. Эти волокна быстро утомляются. Более всего они приспособлены для выполнения кратковременной, но мощной работы (см. главу 27).
Быстрые, устойчивые к утомлению ДЕ (тип II-А). По своим морфофункциональным свойствам этот тип мышечных волокон занимает промежуточные положения между ДЕ I и II- В типов. Это сильные, быстро сокращающиеся волокна, обладающие большой аэробной выносливостью благодаря присущей им возможности использовать для получения энергии как аэробные, так и анаэробные процессы.
У разных людей соотношение числа медленных и быстрых ДЕ в одной и той же мышце определено генетически и может отличаться весьма значительно. Так, например, в четырехглавой мышце бедра человека процент медленных волокон может варьировать от 40 до 98%. Чем больше в мышце процент медленных волокон, тем более она приспособлена к работе на выносливость. И наоборот, лица с
158






 высоким процентом быстрых сильных волокон в большей мере способны к работе, требующей большой силы и скорости сокращения мышц.
высоким процентом быстрых сильных волокон в большей мере способны к работе, требующей большой силы и скорости сокращения мышц.
Регуляция силы сокращения мышц.Для регуляции величины напряжения мышцы центральная нервная система использует три механизма.
1. Регуляция числа активных ДЕ. Чем больше число ДЕ мышцы
включается в работу, тем большее напряжение она развивает. При
необходимости развития небольших усилий и соответственно малой
импульсации со стороны центральных нервных структур, регулиру
ющих произвольные движения, в работу включаются, прежде всего,
медленные ДЕ, мотонейроны, которые имеют наименьший порог
возбуждения. По мере усиления центральной импульсации к работе
подключаются быстрые, устойчивые к утомлению ДЕ, мотонейроны
которых имеют более высокий порог возбуждения. И наконец, при
необходимости увеличения силы сокращения более 20-25 % от
максимальной произвольной силы (МПС), активируются быстрые,
легко утомляемые мышечные волокна, иннервируемые крупными
мотонейронами с самым высоким порогом возбуждения.
Таким образом, первый механизм увеличения силы сокращения состоит в том, что при необходимости повысить величину напряжения мышцы в работу вовлекается большее количество ДЕ. Последовательность включения разных по морфофункиионалъным признакам ДЕ определяется интенсивностью центральных возбуждающих влияний и порогом возбудимости спинальных двигательных нейронов.
2. Регуляция частоты импульсации мотонейронов. При слабых со
кращениях скелетных мышц импульсация мотонейронов составляет
5-10 имп/с. Для каждой отдельной ДЕ чем выше (до определенного
предела) частота возбуждающих импульсов, тем больше сила сокра
щения ее мышечных волокон и тем больше ее вклад в развиваемое
всей мышцей усилие. С увеличением частоты раздражения мотоней
ронов все большее количество ДЕ начинает работать в режиме глад
кого тетануса, увеличивая тем самым свою силу по сравнению с
одиночными сокращениями в 2-3 раза. В реальных условиях мы
шечной деятельности человека большая часть ДЕ активируется в
диапазоне от 0 до 50% МПС. Лишь около 10% ДЕ вовлекаются с
дальнейшим возрастанием силы сокращения. Следовательно, при
увеличении силы сокращения более 50% от максимальной — основ
ное значение, а в диапазоне сил от 75 до 100% МПС — даже
исключительное, принадлежит росту частоты импульсации двигатель
ных нейронов.
3. Синхронизация активности различных ДЕ во времени. При со
кращении мышцы всегда активируется множество составляющих ее
ДЕ. Суммарный механический эффект при этом зависит от того,
как связаны во времени импульсы, посылаемые разными мотоней
ронами к своим мышечным волокнам. При небольших напряжениях
большинство ДЕ работают несинхронно. Совпадение во времени им
пульсов мотонейронов отдельных ДЕ называется синхронизацией.
159




 Чем большее количество ДЕ работает синхронно, тем большую силу развивает мышца.
Чем большее количество ДЕ работает синхронно, тем большую силу развивает мышца.
Синхронизация активности ДЕ играет важную роль в начале любого сокращения и при необходимости выполнения мощных, быстрых сокращений (прыжки, метания и т.п.). Чем больше совпадают периоды сокращения разных ДЕ, тем с большей скоростью нарастает напряжения всей мышцы и тем большей величины достигает амплитуда ее сокращения.
Тонус скелетных мышц.Даже в покое скелетные мышцы редко бывают полностью расслабленными, сохраняя некоторое напряжение, называемое тонусом. Тонус мышц связан с низкочастотной активностью низкопороговых медленных ДЕ, мотонейроны которых активируются влияниями со стороны вышележащих моторных центров и периферических рецепторов (см. главу 15). Тонус мышцы зависит и от ее собственного состояния: эластичности, плотности, условий кровоснабжения, состояния водно-солевого обмена в организме. Человек способен произвольно регулировать тонус своих мышц, особенно после специальной тренировки. Тонус мышц непроизвольно увеличивается после тяжелых физических упражнений, а также, во время психо-эмоционального напряжения.
Кровоснабжение и метаболизм сокращающихся мышц.Между кровоснабжением, метаболизмом и функцией мышечных клеток существует тесная взаимосвязь. Кровоток в скелетных мышцах в покое составляет 2-5 мл • 100 г-1 • мин-1. Поскольку общая масса мышц равна примерно 30 кг, суммарный кровоток через всю мышечную ткань составляет приблизительно 900-1200 мл/мин, то есть 15-20% от минутного объема крови (МОК). При максимальной физической нагрузке мышечный кровоток может достигать 24-27 л/мин при МОК, равном 30 л, то есть 80-90 % общего кровотока.
Величина кровоснабжения сокращающихся мышц зависит от ряда факторов. Во-первых, от степени расширения мышечных сосудов. Благодаря вазодилятации увеличивается радиус всех мышечных сосудов — от артериол до довольно крупных (например, бедренной артерии при сокращении мышц голени). Понижение тонуса гладко-мышечных клеток сосудов во время работы скелетных мышц происходит вследствие действия на них местных метаболических (недостаток О2, избыток СО2, повышенная концентрация ионов водорода, увеличенное содержание АТФ и АДФ, повышенная осмолярность и др.) и физических факторов (уменьшение трансмурального давления, уменьшение степени растяжения гладкомышечных клеток сосудистой стенки, увеличение скорости тока крови в сосуде и др., см. главу 7). Полное раскрытие сосудов происходит через 60-90 с после начала работы. При максимальных ритмических сокращениях небольшой мышечной группы кровоснабжение мышц может достигать 200-250 мл 100 г-1 ■ мин -1.
Во-вторых, от величины механического препятствия кровотоку. При максимальных произвольных сокращениях давление на сосуды
160

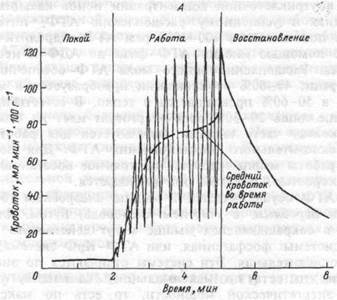
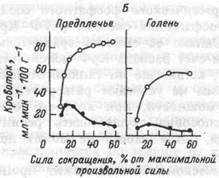
 мышц может в 2- 3 раза превышать уровень систолического артериального давления (АД). Степень сжатия сосудов зависит от силы сокращения. При статической работе кровоток уменьшается уже при нагрузках, превышающих 8- 10% МГТС. При статических усилиях более 30-40 % МПС кровоснабжение мышц практически прекращается. Восстанавливается кровоток через мышцу лишь после окончания работы (рис.4.9.). При ритмических сокращениях кровоток минимален в фазу напряжения и максимален в период расслабления. Средний кровоток при динамической работе всегда намного больше, чем в покое и при статической работе. Это объясняет, почему при динамической работе, при которой сокращение и расслабление постоянно чередуется, мышцы утомляются меньше, чем при статической нагрузке.
мышц может в 2- 3 раза превышать уровень систолического артериального давления (АД). Степень сжатия сосудов зависит от силы сокращения. При статической работе кровоток уменьшается уже при нагрузках, превышающих 8- 10% МГТС. При статических усилиях более 30-40 % МПС кровоснабжение мышц практически прекращается. Восстанавливается кровоток через мышцу лишь после окончания работы (рис.4.9.). При ритмических сокращениях кровоток минимален в фазу напряжения и максимален в период расслабления. Средний кровоток при динамической работе всегда намного больше, чем в покое и при статической работе. Это объясняет, почему при динамической работе, при которой сокращение и расслабление постоянно чередуется, мышцы утомляются меньше, чем при статической нагрузке.

|

|
Рис.4.9. А. Схема изменений кровотока во время ритмических сокращений скелетных мышц человека.
Б. Кровоснабжение мышц предплечья и голени в конце статической работы "до отказа" с различными усилиями (темные кружки) и сразу после ее окончания (светлые кружки).
 161
161



 В-третьих, от размеров работающих мышечных групп. При мощной ритмической работе все сокращающиеся мышцы максимально могут использовать 80-90% МОК. Поэтому, чем меньше по размерам работающие мышечные группы, тем большее количество крови они могут получить в минуту, тем выше будет их работоспособность.
В-третьих, от размеров работающих мышечных групп. При мощной ритмической работе все сокращающиеся мышцы максимально могут использовать 80-90% МОК. Поэтому, чем меньше по размерам работающие мышечные группы, тем большее количество крови они могут получить в минуту, тем выше будет их работоспособность.
В-четвертых, от величины регионарного среднего артериального давления. Чем выше этот показатель при работе, тем больше (при прочих равных условиях) снабжаются кровью активные мышечные волокна. Важное значение в увеличении кровоснабжения ритмически сокращающихся мышц имеет работа "мышечного насоса", способствующего снижению среднего венозного давления и увеличению, благодаря этому, артерио-венозного градиента давления.
Энергетика мышцы.Единственным прямым источником энергии для мышечного сокращения служит АТФ. При активации мышцы повышение внутриклеточной концентрации ионов кальция приводит к сокращению и усиленному расщеплению АТФ, интенсивность метаболизма повышается в 100- 1000 раз. АТФ гидролитически расщепляется с помощью миозин-АТФ-фазы до АДФ и неорганического фосфата. Расщепление одного моля АТФ обеспечивает около 48 кДж энергии. 40- 50% этой энергии преобразуется в механическую работу, а 50-60% превращается в тепло. В естественных условиях в мышце лишь 20- 30% всех энерготрат идет на механическую работу, поскольку часть энергии используется для работы ионных насосов и окислительного восстановления АТФ. Для поддержания длительной работы мышц требуется постоянное восстановление АТФ с той же скоростью, с какой она расходуется.
Ресинтез АТФ осуществляется в мышце анаэробным (без участия кислорода) и аэробным (с участием кислорода) путем. Для образования АТФв сокращающейся мышце могут действовать три энергетические системы: фосфагенная, или АТФ-КрФ система, гликоли-тическая и окислительная. Эти системы отличаются по энергетической емкости, то есть по максимальному количеству образуемой энергии и энергетической мощности, то есть по максимальному количеству энергии, образующейся в единицу времени.
Восстановление АТФ во время сокращения осуществляется почти мгновенно, как только она расщепляется до АДФ. Происходит это за счет энергии другого высокоэнергетического фосфатного соединения — креатинфосфата (КрФ). Фосфагенная система (АТФ-КрФ) обладает наибольшей мощностью. Однако ее емкость сравнительно невелика. Поэтому, ресинтез АТФ за счет распада КрФ используется лишь в самом начале любой работы, когда еще ни гликолитическая, ни, тем более, окислительная системы не успевают развернуться, а так же при работе максимальной мощности, при которой только анаэробная фосфагенная система способна осуществлять ресинтез АТФ с подобной скоростью. Правда, такая работа не может продолжаться более 5-6 с.
В основе деятельности гликолитической системы лежит процесс анаэробного расщепления углеводов (гликогена, глюкозы) до молоч-
162

 ной кислоты. При этом каждая молекула глюкозы дает энергию для образования 3 молекул АТФ. Во время этой реакции скорость образования АТФ в 2-3 раза выше, а механическая работа в 2-3 раза больше, чем при длительной аэробной работе. Однако, емкость гликолитической системы намного меньше, чем окислительной (хотя в 2,5 раза больше, чем фосфагенной). Поэтому работа при анаэробном гликолизе в качестве ведущего источника энергии может продолжаться лишь короткое время — от 20 с до 1-2 мин. Заканчивается такая работа на фоне значительного накопления молочной кислоты в клетках и в крови.
ной кислоты. При этом каждая молекула глюкозы дает энергию для образования 3 молекул АТФ. Во время этой реакции скорость образования АТФ в 2-3 раза выше, а механическая работа в 2-3 раза больше, чем при длительной аэробной работе. Однако, емкость гликолитической системы намного меньше, чем окислительной (хотя в 2,5 раза больше, чем фосфагенной). Поэтому работа при анаэробном гликолизе в качестве ведущего источника энергии может продолжаться лишь короткое время — от 20 с до 1-2 мин. Заканчивается такая работа на фоне значительного накопления молочной кислоты в клетках и в крови.
Таким образом, гликолитическая энергетическая система функционирует в том случае, когда сокращающиеся мышцы снабжаются кислородом в недостаточной степени. По мере удлинения времени работы и развертывания кислородтранспортной системы (на это уходит 2-3 мин), ресинтез АТФ начинает осуществляться, в основном, за счет окислительного фосфорилирования.
При продолжительной и мало интенсивной работе потребности мышц в кислороде удовлетворяются полностью. Необходимая для ресинтеза АТФ энергия поступает в результате окисления углеводов и жиров. Чем больше мощность работы, тем относительно меньше энергетический вклад окисляемых жиров в общую энергопродукцию сокращающихся мышц.
При работе очень большой мощности окисляются, в основном, углеводы, а при малоинтенсивной — жиры. При аэробном расщеплении одной молекулы глюкозы образуется 38 молекул АТФ, то есть значительно больше, чем в процессе анаэробного гликолиза. Емкость окислительной энергетической системы в тысячи раз превышает емкость фосфагенной и гликолитической систем. Именно поэтому, при достаточном кровоснабжении мышечных волокон и поступлении к ним кислорода, работа может выполняться мышцами на протяжении многих часов.
Дата добавления: 2016-03-27; просмотров: 2571;