МЕХАНИЗМ ПЕРИОДИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ДЫХАТЕЛЬНОГО ЦЕНТРА
Рецепторы легких, дыхательных мышц, верхних дыхательных путей раздражаются дыхательными движениями, однако дыхательный центр ствола мозга способен обеспечить смену дыхательных фаз за счет деятельности своих внутренних механизмов.
При введении животному миорелаксантов (вещества, превращающие передачу возбуждения с двигательных нервных волокон на мышцы) наступал полный паралич скелетной мускулатуры, в том числе и дыхательной. Животных переводили на искусственную вентиляцию легких, которую через некоторое время прекращали. Смена фаз инспирации и экспирации продолжалась и в этих условиях, о чем свидетельствовало периодическое возбуждение диафрагмальных мотонейронов и других дыхательных нейронов.
Таким образом, дыхательный центр содержит механизмы, необходимые для формирования спонтанного периодического возбуждения своих нейронов,и в этом смысле обладает автоматизмом.
Автоматизм дыхательного центра отличается рядом признаков от автоматизма водителя сердечного ритма. 1. Периодическое возбуждение дыхательного центра обусловлено взаимодействием многих нервных клеток, среди которых особо важную роль играют тормозные нейроны. 2. Для автоматической деятельности .дыхательного центра необходимо постоянное (тоническое) поступление к нему сигналов, повышающих возбудимость дыхательных нейронов, от хеморецепторов, а также от ретикулярной формации ствола мозга. 3. Автоматическая деятельность дыхательного центра человека находится под сильно выраженным произвольным контролем. Человек может в широких пределах изменять частоту и глубину своего дыхания.
Теории возникновения периодической деятельности дыхательного центра.Механизмы периодической деятельности дыхательного центра полностью еще не выяснены. В частности,.несмотря на продолжающиеся исследования, нет достаточных сведений о связях между группами (популяциями) дыхательных нейронов. Неизвестна природа торможения инспираторных нейронов при смене вдоха на выдох.
Рассмотрим модель механизма периодической деятельности дыхательного центра. В этой модели обобщены многие экспериментальные данные, и она является предметом современных исследований.
Основа модели—два нейронных механизма: 1) генератор центрального инспира-торного возбуждения (ЦИВ) и 2) механизм выключения инспирации.
Генератор ЦИВ обеспечивает возникновение и постепенное усиление возбуждения инспираторных мышц по ходу вдоха. Он представляет собой совокупность инспираторных нейронов 1а, тормозящихся при раздражении рецепторов растяжения легких. Условием возникновения ЦИВ является достаточно интенсивный поток возбуждающих сигналов от центральных и периферических хеморецепторов (рис. 161). От стимуляции хеморецепторов и, следовательно, от газового состава крови зависит скорость нарастания инспира-торной активности по ходу вдоха: чем больше активируются хеморецепторы, тем больше скорость нарастания ЦИВ.
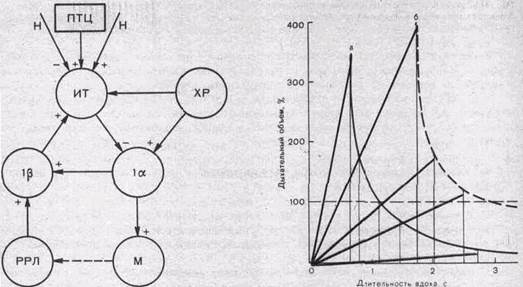
Рис.161. Модель механизма смены дыхательных фаз.
ХР — хеморецепторы; 1а — генератор центрального инспираторного возбуждения; 1р — инспираторные нейроны, на которых конвергируют импульсы нейронов 1а и рецепторов растяжения легких (РРЛ); ИТ— инспираторно-тормозящие нейроны; М — мотонейроны дыхательных мышц и сами мышцы; ПТЦ — пневмо-таксический центр; Н — нисходящие влияния на бульбарный дыхательный центр; + — возбуждающие связи; — — тормозные связи.
Рис.162. Зависимость длительности вдохов от их глубины.
За 100 % принят дыхательный объем при эипноэ. Прямые, начинающиеся от нуля,— скорость развития вдохов до их перерыва (точки пересечения с кривыми), а — до перерезки блуждающих нервов, б — после перерезки блуждающих нервов.
У животного при помощи гипервентиляции вызывали гипокапническое апноэ. Затем гипервен-тиляцию прекращали. Появлялось редкое поверхностное дыхание. По мере накопления двуокиси углерода в крови глубина дыхания увеличивалась. Добавляя во вдыхаемый воздух двуокись углерода, наблюдали увеличение глубины дыхания до максимальных величин. Это можно представить графически (рис. 162). Величины дыхательного объема, пропорциональные ЦИВ, отложены на оси ординат, а на абсциссе—длительность вдохов (t|). Прямые от начала координат отражают развитие во времени ЦИВ.
Основой механизма торможения инспирации считают инспираторные нейроны 1(3, возбуждаемые афферентными сигналами от рецепторов растяжения легких. Когда возбуждение нейронов 1р достигает определенного порогового уровня, возбуждение инспи-раторных нейронов резко ослабевает или прекращается, т. е. вдох сменяется выдохом. Одновременно экспираторные нейроны вентрального дыхательного ядра освобождаются от тормозящего влияния нейронов 1а (см. рис. 161). Возбуждение экспираторных нейронов, обусловленное сигналами от хеморецепторов, усиливается по ходу выдоха.
На рис. 162 моменты выключения инспираций соединены кривой (так называемая кривая порогов выключения инспираций). Кривая близка к гиперболе. Чем сильнее раздражение хеморецепторов, тем больше скорость развития ЦИВ, меньше длительность инспираций и больше дыхательный объем.

|
Рис.163. Изменения дыхания после перерезки блуждающих нервов и разрушения пневмо-таксического центра.
а — нормальное дыхание — эйпнчэ; б — после перерезки блуждающих нервов и разрушения пнев-мотаксического центра вдохи стали необычно продолжительными (апнейзисы); в—отметка времени 10 с.
Примечательно, что влияние сигналов or рецепторов растяжения легких до достижения порога возбуждения нейронов 1(3 не сказывается на скорости развития ЦИВ. После перерезки блуждающих нервов, выключающей влияния рецепторов растяжения легких, скорость увеличения ЦИВ по ходу вдоха при данном газовом составе крови не изменяется. Значит, инспираторная активность тормозится скачком (по принципу «все или ничего»).
Установлено, что нейроны 1(3 не оказывают непосредственного тормозящего влияния на нейроны 1а. Поэтому предполагают существование инспираторно-тормозящих нейронов. Какие нейроны выполняют эту функцию, точно неизвестно. Предполагается, что ими могут быть инспираторно-экспираторные или «ранние» экспираторные нейроны. Чтобы объяснить увеличение дыхательного объема при усилении раздражения хеморе-цепторов, сделано предположение, что сигналы от хеморецепторов не только повышают возбуждение нейронов 1«, но и снижают возбудимость инспираторно-тормозящих нейронов,
Во многих условиях период выдоха определяется длительностью предшествующего вдоха, причем эта зависимость носит линейный характер. При данном газовом составе крови преждевременное выключение инспирации (например, коротким раздражением рецепторов растяжения легких) приводит к пропорциональному укорочению экспирации. Предполагают, что возбуждение инспираторно-тормозящих нейронов после выключения инспирации прекращается не сразу, а затухает постепенно. Генератор ЦИВ постепенно освобождается от торможения. Когда возбуждение инспираторно-тормозящих нейронов снижается до определенного уровня, возникает очередная инспирация.
При развитии гиперпноэ длительность выдохов уменьшается за счет двух факторов:
редкой импульсации рецепторов растяжения легких и усиления раздражения ирритант-ных рецепторов. Оба фактора способствуют более раннему наступлению следующей инспирации.
Рассмотренные нейронные механизмы (генератор ЦИВ, механизм выключения вдоха, экспираторные нейроны) находятся в продолговатом мозге, они входят в состав дорсальных и вентральных дыхательных ядер. Эти механизмы испытывают постоянные нисходящие влияния, в частности от пневмотаксического центра моста.
Пневмотаксический центр. Поперечная перерезка моста ниже четверохолмия сопровождается изменениями дыхания, сходными с теми, которые наступают после перерезки блуждающих нервов. Частота дыхания снижается вследствие увеличения продолжительности как вдохов, так и выдохов. Если дополнительно произвести перерезку блуждающих нервов, вдохи становятся необычайно длительными, они продолжаются десятки секунд или минуты. Долго непрерывающиеся вдохи называются апнейяисами (рис. 163). Структуры передней части моста, разрушение которых после перерезки блуждающих нервов ведет к возникновению апнейзисов, получили название .пневмотаксического центра.
Установлены двусторонние связи бульбарного дыхательного и пневмотаксического центров. При раздражении отдельных зон пневмотаксического центра может быть преждевременно вызван вдох или выдох.
Таким образом, в передней части моста находится центральный механизм, которыи, подобно сигналам от рецепторов легких, способствует смене дыхательных фаз, т. е. периодической деятельности дыхательного центра. Пневмотаксический центр увеличивает скорость развития инспираторной активности и повышает возбудимость механизма выключения вдоха. Помимо этого, пневмотаксический центр ускоряет наступление следующей инспирации.
Пневмотаксический центр не является генератором дыхательной периодики, но он постоянно участвует в регуляции частоты дыхания.
На периодической деятельности дыхательного центра сказывается активность ретикулярной формации средних и нижних областей моста. После разрушения этих областей остановки дыхания нс возникают. У бульбарных животных (после перерезки мозга между мостом и продолговатым мозгом) обычно наблюдается судорожное дыхание, или гаспинг. Для него характерны короткие быстрые вдохи и длительные (10—20 с) экспираторные паузы. Реже отмечается дыхание, напоминающее эйпноэ, однако ритм такого дыхания неравномерен, а вентиляция легких недостаточна для поддержания газового состава крови на нормальном уровне.
Влияние на дыхательный центр раздражения различных рецепторов и отделов ЦНС;
Дата добавления: 2016-03-27; просмотров: 4395;