Роль хеморецепторов в регуляции дыхания
Давно установлено, что деятельность дыхательного центра зависит от состава крови, поступающей в мозг по общим сонным артериям.
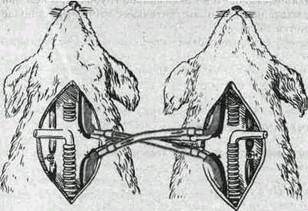
Это было показано Фредериком (1890) в опытах с перекрестным кровообращением. У двух собак, находившихся под наркозом, перерезали и соединяли перекрестие сонные артерии и отдельно яремные вены' (рис. 158). После такого соединения и перевязки позвоночных артерий голова первой собаки снабжалась кровью второй собаки, голова второй собаки — кровью первой. Если у одной из собак, например у первой, перекрывали трахею и вызывали таким путем асфиксию, то гиперпноэ развивалось у второй собаки. У первой же собаки, несмотря на увеличение в артериальной крови напряжения двуокиси углерода и снижение напряжения кислорода, через некоторое время наступало апноэ. Это объясняется тем, что в сонную артерию первой собаки поступала кровь второй собаки, у которой в результате гипервентиляции в артериальной крови снижалось напряжение двуокиси углерода.
Двуокись углерода, водородные ионы и умеренная гипоксия вызывают усиление дыхания, действуя не непосредственно на нейроны дыхательного центра. Возбудимость дыхательных нейронов, как и других нервных клеток, под влиянием этих факторов снижается. Следовательно, эти факторы усиливают деятельность дыхательного центра, оказывая влияние на специальные хеморецепторы. Имеется две группы хеморецепторов, регулирующих дыхание: периферические (артериальные) и центральные (медуллярные).
Артериальные хеморецепторы. Хеморецепторы, стимулируемые увеличением напряжения двуокиси углерода и снижением напряжения кислорода, находятся в каротидных синусах и дуге аорты. Они расположены в специальных маленьких тельцах, обильно снабжаемых артериальной кровью. Важными для регуляции дыхания являются каротид-ные хеморецепторы. Аортальные хеморецепторы на дыхание влияют слабо и имеют большее значение для регуляции кровообращения.
Каротидные тельца расположены в развилке общей сонной артерии на внутреннюю и наружную. Масса каждого каротидного тельца всего около 2 мг. В нем содержатся относительно •крупные эпителиоидные клетки I типа, окруженные мелкими интерстициальными клетками II типа. С клетками I типа контактируют окончания афферентных волокон синусного нерва (нерва Геринга), который является ветвью языкоглоточного нерва. Какие структуры тельца — клетки I или II типа либо нервные волокна — являются собственно рецепторами, точно не установлено.
Хеморецепторы каротидных и аортальных телец являются уникальными рецептор-ными образованиями, на которые гипоксия оказывает стимулирующее влияние. Афферентные сигналы в волокнах, отходящих от каротидных телец, можно зарегистрировать и при нормальном (100 мм рт. ст.) напряжении кислорода в артериальной крови. При снижении напряжения кислорода от 80 до 20 мм рт. ст. частота импульсов увеличивается особенно значительно.
Кроме того, афферентные влияния каротидных телец усиливаются при повышении в артериальной крови напряжения двуокиси углерода и концентрации водородных ионов. Стимулирующее действие гипоксии и гиперкапнии на данные хеморецепторы взаимно усиливается. Наоборот, в условиях гипероксии чувствительность хеморецепторов к двуокиси углерода резко снижается.
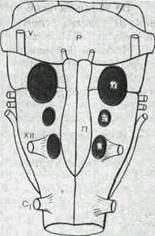
| Рис.159. Расположение хемо-рецепторов на вентральной по ве р хно сти п родол го ва то го мозга. М, L, S — поля. участвующие в хемореиепции; Р — мост; П — пирамида; V и XII — черепно-мозговые нервы; С] — первый сп и и ном оз гов о и корешок. |
 |
 Хеморецепторы телец особенно чувствительны к колебаниям газового состава крови. Степень их активации возрастает при колебаниях напряжения кислорода и двуокиси
Хеморецепторы телец особенно чувствительны к колебаниям газового состава крови. Степень их активации возрастает при колебаниях напряжения кислорода и двуокиси
Рис. 158. Схема опыта Фредерика с перекрестным кровообращением.
углерода в артериальной крови даже в зависимости от фаз вдоха и выдоха при глубоком и редком дыхании.
Чувствительность хеморецепторов находится под нервным контролем. Раздражение эфферентных парасимпатических волокон снижает чувствительность, а раздражение симпатических волокон повышает ее.
Хеморецепторы (особенно каротидных телец) информируют дыхательный центр о напряжении кислорода и двуокиси углерода в крови, направляющейся к мозгу.
Центральные хеморецепторы. После денервации каротидных и аортальных телец исключается усиление дыхания в ответ на гипоксию. В этих условиях гипоксия вызывает только снижение вентиляции легких, но зависимость деятельности дыхательного центра от напряжения двуокиси углерода сохраняется. Она обусловлена функцией центральных хеморецепторов.
Центральные хеморецепторы были обнаружены в продолговатом мозге латеральное пирамид (рис. 159). Перфузия этой области мозга раствором со сниженным рН резко усиливает дыхание. Если рН раствора увеличить, то дыхание ослабевает (у животных с денервированными каротидными тельцами останавливается на выдохе, наступает апноэ). То же присходит при охлаждении или обработке местными анестетиками этой поверхности продолговатого мозга.
Хеморецепторы расположены в тонком слое мозгового вещества на глубине не более 0,2 мм. Обнаружены два рецептивных поля, обозначаемые буквам М и L. Между ними находится небольшое поле S. Оно нечувствительно к концентрации ионов Н4', но при его разрушении исчезают эффекты возбуждения полей М и L. Вероятно, здесь проходят афферентные пути от сосудистых хеморецепторов к дыхательному центру.
В обычных условиях рецепторы продолговатого мозга постоянно стимулируются ионами Н4', находящимися в спинномозговой жидкости. Концентрация Н'1' в ней зависит от напряжения двуокиси углерода в артериальной крови, она увеличивается при гиперкапнии.
Центральные хеморецепторы оказывают более сильное влияние на деятельность дыхательного центра, чем периферические. Они существенно изменяют вентиляцию легких. Так, снижение рН спинномозговой жидкости на 0,01 сопровождается увеличением вентиляции легких на 4 л/мин. Вместе с тем центральные хеморецепторы реагируют на изменение напряжения двуокиси углерода в артериальной крови позже (через 20—30 с), чем периферические хеморецепторы (через 3—5 с). Указанная особенность обусловлена тем, что для диффузии стимулирующих факторов из крови в спинномозговую жидкость и далее в ткань мозга необходимо время.
Сигналы, поступающие от центральных и периферических хеморецепторов, являются необходимым условием периодической активности дыхательного центра и соответствия вентиляции легких газовому составу крови. Импульсы от центральных хеморецепторов усиливают возбуждение как инспираторных, так и экспираторных нейронов дыхательного центра продолговатого мозга.
Дата добавления: 2016-03-27; просмотров: 4645;