Транспорт двуокиси углерода
Двуокись углерода переносится кровью в трех формах. Из венозной крови можно извлечь около 58 об. % (580 мл/л) двуокиси углерода, из них лишь около 2,5 об. % находятся в состоянии физического растворения. Остальное количество двуокиси углерода химически связано и содержится в виде кислых солей угольной кислоты {51 об. %) и карбгемоглобина (4,5 об. %}.
Двуокись углерода непрерывно образуется в клетках и диффундирует в кровь тканевых капилляров. В эритроцитах она соединяется с водой и образует угольную кислоту. Этот процесс катализируется (ускоряется в 20000 раз) ферментом карбоангидразой. Карбоангидраза содержится в эритроцитах, в плазме крови ее нет. Поэтому гидратация двуокиси углерода происходит практически только в эритроцитах. В зависимости от напряжения двуокиси углерода карбоангидраза катализирует как образование угольной кислоты, так и расщепление ее на двуокись углерода и воду (в капиллярах легких).
Часть молекул двуокиси углерода соединяется в эритроцитах с гемоглобином, образуя карбгемоглобин.
Благодаря указанным процессам связывания напряжение двуокиси углерода в эритроцитах оказывается невысоким. Поэтому все новые количества двуокиси углерода диффундируют внутрь эритроцитов. Концентрация ионов НСОз", образующихся при диссоциации солей угольной кислоты, в эритроцитах возрастает. Мембрана эритроцитов обладает высокой проницаемостью для анионов. Поэтому часть ионов НСОГ поступает в плазму крови. Взамен ионов НСОГ в эритроциты из плазмы входят ионы С1~, отрицательные заряды которых уравновешиваются ионами К"1'. В плазме крови увеличивается количество бикарбоната натрия (NaHCOa).
Накопление ионов внутри эритроцитов сопровождается повышением в них осмотического давления. Поэтому объем эритроцитов в капиллярах большого круга кровообращения несколько увеличивается.
Для связывания большей части двуокиси углерода исключительно большое значение имеют свойства гемоглобина как кислоты. Оксигемоглобин имеет константу диссоциации в 70 раз большую, чем дезоксигемоглобчн. Оксигемоглобин — более сильная кислота, чем угольная, а дезоксигемоглобин — более слабая. Поэтому в артериальной крови оксигемоглобин, вытеснивший ионы К'1' из бикарбонатов, переносится в виде соли КНЬОа. В тканевых капиллярах часть КНЬОг отдает кислород и превращается в КНЬ. Из него угольная кислота как более сильная вытесняет ионы К4':
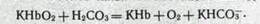
Таким образом, превращение оксигемоглобина в гемоглобин сопровождается увеличением способности крови связывать двуокись углерода. Это явление носит название эффекта Холдейна. Гемоглобин служит источником катионов (К^), необходимых для связывания угольной кислоты в форме бикарбонатов.
Итак, в эритроцитах тканевых капилляров образуется дополнительное количество бикарбоната калия, а также карбгемоглобин, а в плазме крови увеличивается количество бикарбоната натрия. В таком виде двуокись углерода переносится к легким.
В капиллярах малого круга кровообращения напряжение двуокиси углерода снижается. От карбгемоглобина отщепляется СОг. Одновременно происходит образование оксигемоглобина, увеличивается его диссоциация. Оксигемоглобин вытесняет калий из бикарбонатов. Угольная кислота в эритроцитах (в присутствии карбоангидразы) быстро разлагается на НаО и С02. Ионы НСОз входят в эритроциты, а ионы С1~ выходят в плазму крови, где уменьшается количество бикарбоната натрия. Двуокись углерода диффундирует в альвеолярный воздух. Схематически все эти процессы представлены на рис. 153.
ОБМЕН ГАЗОВ В ТКАНЯХ
Наименьшее напряжение кислорода наблюдается в местах его потребления — митохондриях клеток, в которых кислород используется для процессов биологического окисления. Молекулы кислорода, освобождающиеся по ходу кровеносных капилляров в результате диссоциации оксигемоглобина, диффундируют в направлении более низких величин напряжения кислорода. Напряжение кислорода в тканях зависит от многих факторов: скорости тока крови, геометрии капилляров и расстояния между ними, расположения клеток по отношению к капиллярам, интенсивности окислительных процессов и т. д. В тканевой жидкости около капилляров напряжение кислорода значительно ниже (20—40 мм рт. ст.), чем в крови. Особенно низко оно в уч-астках тканей, равноудаленных от соседних капилляров. При большой интенсивности окислительных процессов напряжение кислорода в клетках может приближаться к нулю. Увеличение скорости кро-вотока резко повышает напряжение кислорода в тканях. Например, увеличение скорости тока крови вдвое может повысить напряжение кислорода в нервной клетке на 10 мм рт. ст. В мышцах увеличению снабжения кислородом способствует раскрытие так называемых резервных капилляров.
Наибольшее напряжение двуокиси углерода (до 60 мм рт. ст.) отмечается в клетках в результате образования этого газа в митохондриях. В тканевой жидкости напряжение двуокиси углерода изменчиво (в среднем 46 мм рт. ст.), а в артериальной крови составляет 40 мм рт. ст. Двуокись углерода диффундирует по градиенту напряжений в кровеносные капилляры и транспортируется кровью к легким.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Вентиляция легких осуществляется возвратно-поступательным движением воздуха в результате периодических сокращений дыхательных мышц. Частота, сила и форма этих сокращений соответствуют потребностям организма.
Иннервация дыхательных мышц. Подобно другим скелетным мышцам, дыхательные мышцы иннервируются соматическими нервными волокнами. Если перерезать нервы, подходящие к дыхательным мышцам, последние оказываются парализованными. Например, перерезка диафрагмального нерва ведет к прекращению сокращений соответствую-. щей половины диафрагмы. Значит, периодические сокращения дыхательных мышц вызываются импульсами, поступающими из мозга.
Мотонейроны, аксоны которых иннервируют диафрагму, находятся в спинном мозге в передних рогах серого вещества/// и IV шейных сегментов. Мотонейроны межреберных мышц и мышц живота расположены в грудных сегментах спинного мозга. Вместе с интернейронами, участвующими в координации сокращений, мотонейроны образуют спинномозговые центры дыхания (ядра дыхательных мышц).
После отделения головного мозга от спинного на уровне верхних шейных сегментов дыхательные движения прекращаются. Лишь изредка удается наблюдать слабые сокращения дыхательных мышц, но они имеют неправильные ритм и форму. Если перерезать мозг на уровне нижних шейных сегментов, дыхательная активность диафрагмы сохраняется, а межреберных мышц — прекращается. Следовательно, в регуляции дыхания принимают участие и центры головного мозга.
ДЫХАТЕЛЬНЫЙ ЦЕНТР
После перерезки мозгового ствола между средним мозгом и мостом (децеребрация) дыхание у животных в состоянии покоя существенно не нарушается. Значит, центральные механизмы, управляющие дыхательными движениями, находятся в продолговатом мозге и мосту. Совокупность сгруппированных здесь нейронов носит название бульбопонтин-ного дыхательного центра. После отделения моста от продолговатого мозга дыхательный ритм может сохраниться, но будет отличаться от нормального. Следовательно, важнейшие структуры дыхательного центра находятся в продолговатом мозге. Это бульварный дыхательный центр. Его разрушение локальным повреждением полностью прекращает периодические сокращения дыхательных мышц.
Дыхательный цикл. Явления, происходящие в аппарате внешнего дыхания между началом следующих друг за другом вдохов, называются дыхательным циклом. Его длительность у человека составляет от 3 до 5 с. Все уровни дыхательного центра обеспечивают характерный рисунок (паттерн) возбуждения дыхательных мышц.
В определенный момент возникает возбуждение диафрагмальных мотонейронов (начало фазы инспирации). Это возбуждение постепенно усиливается (рис. 154) за счет увеличения частоты разрядов отдельных мотонейронов, а также вследствие вовлечения в возбуждение новых («поздних») мотонейронов (в диафрагмаль-ном нерве содержится около 1000 аксонов мотонейронов). При спокойном дыхании у человека возбуждение усиливается в течение 1—2,5 с. В результате сила сокращения диафрагмы постепенно возрастает. Затем возбуждение диафрагмальных мотонейронов резко ослабевает: инспирация сменяется фазой экспирации. Через 2—3,5 с наступает следующая инспирация. Как правило, длительность инспирации меньше, чем экспирации.
Возбуждение инспираторных межреберных мышц имеет рисунок возбуждения, близкий к возбуждению диафрагмы, но обычно возникает несколько позже, чем возбуждение диафрагмы. 
Рис.154. Форма инспираторной и экспираторной активности. 1 — потенциалы действия одиночного двигательного волокна диафрагмального нерва; 2 — интегрированные потенциалы действия диафрагмального нерва; 3 — интегрированные потенциалы действия экспираторной мышцы; 4 — отметка времени I с.
Рис.155. Потенциалы действия одиночных инспираторного (1) и экспираторного (2) нейронов. 3—интегрированные потенциалы действия диафрагмального нерва; 4 — фазы дыхательного цикла: И-инспирация; Э — экспирация.
При активном выдохе возбуждение мышц живота и внутренних межреберных мышц усиливается по ходу экспирации и резко ослабевает перед началом следующей инспирации (см. рис. 154).
Дыхательные нейроны. Для установления локализации дыхательного центра использовали методы разрушения и раздражения ограниченных участков мозга. Однако основные сведения о расположении структур дыхательного центра были получены при помощи микроэлектродов путем регистрации потенциалов действия отдельных нейронов, возбуждающихся в соответствии с фазами дыхательного цикла.
Обнаружены две основные группы дыхательных нейронов — инспираторные и экспираторные. Потенциалы действия типичных инспираторных нейронов возникают за 0,1— 0,2 с до начала вдоха. При вдохе частота разрядов постепенно увеличивается и к концу инспирации достигает 70—100 в 1 с (при сильных вдохах до 300). При смене вдоха выдохом разряды прекращаются или их частота резко уменьшается.
Частота потенциалов действия экспираторных нейронов увеличивается в течение выдоха. Прекращение разрядов или уменьшение их частоты происходит перед самым началом следующей инспирации (рис. 155).
Реже встречаются инспираторные и экспираторные нейроны, максимальная частота разрядов которых соответствует началу данной фазы дыхательного цикла («ранние» нейроны) или моменту смены дыхательных фаз (инспираторно-экспираторные и экспираторно-инспираторные нейроны).
Локализация дыхательных нейронов. В правой и левой половинах продолговатого мозга содержатся по два скопления дыхательных нейронов — дорсальные и вентральные дыхательные ядра. Ориентиром их расположения служит задвижка (обекс), находящаяся у нижнего угла ромбовидной ямки.
Дорсальное дыхательное ядро входит в состав серого вещества, окружающего одиночный пучок (ядро одиночного пучка) (рис. 156). Оно содержит преимущественно инспираторные нейроны, аксоны которых направляются в основном к диафрагмальным ядрам шейного отдела спинного мозга. Коллатерали аксонов следуют также в вентральное дыхательное ядро, образуя возбуждающие синапсы ня инспираторных нейронах. Таким образом, возбуждение нейронов дорсального дыхательного ядра тормозит возбуждение экспираторных нейронов вентрального дыхательного ядра. Экспираторные нейроны в дорсальном дыхательн-ом ядре встречаются редко (их здесь около 5%).
Вентральное дыхательное ядро имеет большую протяженность — от каудального края ядра лицевого нерва до I шейного сегмента спинного мозга. Оно включает в<'ебя обоюдное ядро, в котором находятся мотонейроны мышц гортани и глотки. Часть вентрального ядра, расположенная латеральное и каудальнее обоюдного ядра, называется
|
|
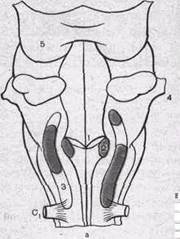
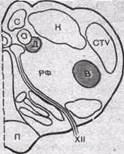
Рис.156. Дыхательные ядра продолговатого мозга. а — проекция дыхательных ядер на дорсальную поверхность продолговатого мозга: 1 — обекс; 2 — дорсальное дыхательное ядро; 3 — вентральное дыхательное ядро; 4 — граница моста и продолговатого мозга; 5 — заднее двухолмие; Ci — корешок спинного мозга. Слева — области скопления инспираторных нейронов, справа — области скопления экспираторных нейронов; б — схема поперечного среза продолго-
.атого мозга на уровне obex; Д — дорсальное дыхательное ядро; В — вентральное дыхательное ядро; С — стройное ядро; К — клиновидное ядро; О—одиночный пучок; CTV — ядро спинального тракта тройничного нерва; РФ — ретикулярная формация, XII — подъязычный нерв П — пирамида.
ретроамбигуальным ядром. Вентральное дыхательное ядро расположено в вентролате-ральной области продолговатого мозга. В вентральном дыхательном ядре содержатся как инспираторные, так и экспираторные нейроны.
Большая часть нейронов вентрального дыхательного ядра посылает аксоны к спинномозговым ядрам дыхательных мышц, в основном межреберных мышц и мышц живота. Примерно 25% волокон разветвляется в области диафрагмальных ядер, 90% аксонов инспираторных нейронов и все аксоны экспираторных нейронов перекрещиваются в продолговатом мозге и следуют к спинномозговым ядрам в вентральном канатике и передней части бокового канатика белого вещества противоположной стороны спинного мозга. Кроме того, в вентральном дыхательном ядре имеются нейроны, аксоны которых оканчиваются в продолговатом мозге (проприобульбарные нейроны).
Небольшое количество дыхательных нейронов встречается и вне дыхательных ядер — в ретикулярной формации продолговатого мозга и моста.

|
|
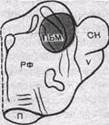
Третье компактное скопление дыхательных нейронов было обнаружено у животных после перерезки блуждающих нервов в передней части моста, сразу за четверохолмием. Это скопление находится в медиальном парабрахиальном ядре (латеральнее его). При сохраненных блуждающих нервах нейроны этих ядер имеют непрерывную тоническую импульсную активность. Дыхательно? ядро переднего моста носит название пневмотак-сического центра (рис. 157). Таким образом, дыхательный центр имеет сложную нейронную структуру.
Рис. 157. Локализация пневмо-таксического центра. а — проекция пневмотаксического центра на дорсальную поверхность моста; б — схема поперечного среза через переднюю часть моста. Выделен пневмотаксический центр. ПБМ — медиальное парабрахиаль-ное ядро; СН — средние ножки мозжечка; V — тройничный нерв;
П — пирамида-
Дата добавления: 2016-03-27; просмотров: 1306;