Транспорт кислорода
В 100 мл крови при температуре тела растворяется лишь 0,3 мл кислорода. Кислород, растворяющийся в плазме крови капилляров малого круга кровообращения, диффундирует в эритроциты, сразу же связывается гемоглобином, образуя оксигемоглобин, в котором кислорода 190 мл/л. Скорость связывания кислорода велика: время полунасыщения гемоглобина кислородом около 3 мс. В капиллярах альвеол с соответствующими вентиляцией и перфузией практически весь гемоглобин превращается в оксигемоглобин.
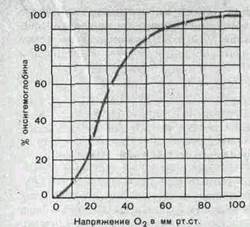
Кривая диссоциации оксигемоглобина. Превращение гемоглобина в оксигемоглобин определяется напряжением растворенного кислорода. Графически эта зависимость выражается кривой диссоциации оксигемоглобина (рис. 151).
Когда напряжение кислорода равно нулю, в крови находится только восстановленный гемоглобин (дезоксигемоглобин). Повышение напряжения кислорода сопровождается увеличением количества оксигемоглобина. Но данная зависимость существенно отличается от линейной, кривая имеет S-образную форму. Особенно быстро (до 75%) уровень оксигемоглобина возрастает при увеличении напряжения кислорода от 10 до 40 мм рт. ст. При 60 мм рт. ст. насыщение гемоглобина кислородом достигает 90%, а при дальнейшем повышении напряжения кислорода приближается к полному насыщению очень медленно. Таким образом, кривая диссоциации оксигемоглобина состоит из двух основных частей — крутой и отлогой.
Отлогая часть кривой, соответствующая высоким (более 60 мм.рт. ст.) напряжениям кислорода, свидетельствует о том, что в этих условиях содержание оксигемоглобина лишь слабо зависит от напряжения кислорода и его парциального давления во вдыхаемом и альвеолярном воздухе. Так, подъем на высоту 2 км над уровнем моря сопровождается снижением атмосферного давления с 760 до 600 мм рт. ст., парциального давления кислорода в альвеолярном воздухе со 105 до 70 мм рт. ст., а содержание оксигемоглобина снижается лишь на 3%. Таким образом, верхняя отлогая часть кривой диссоциации отражает способность гемоглобина связывать большие количества кислорода несмотря
|
Рис.151. Кривая диссоциации оксигемо-глобина при напряжении двуокиси углерода 40 мм рт ст удумеренное снижение его парциального давления во вдыхаемом воздухе. И в этих условиях ткани достаточно снабжаются кислородом.
Крутая часть кривой диссоциации соответствует напряжениям кислорода, обычным для тканей организма (35 мм рт. ст. и ниже). В тканях, поглощающих много кислорода (работающие мышцы, печень, почки), оксигемоглобин диссоциирует в большей степени, иногда почти полностью. В тканях, в которых интенсивность окислительных процессов мала, большая часть оксигемоглобина не диссоциирует. Переход тканей из состояния покоя в деятельное состояние (сокращение мышц, секреция желез) автоматически создает условия для увеличения диссоциации оксигемоглобина и увеличения снабжения тканей .кислородом.
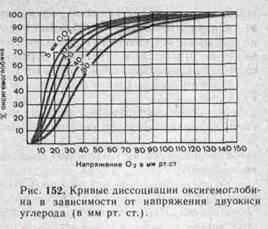
Сродство гемоглобина к кислороду (отражается кривой диссоциации оксигемоглобина) непостоянно. Особенно значительно на него влияют следующие факторы. 1. В эритроцитах содержится особое вещество 2, 3-дифосфоглицерат. Его количество увеличивается, в частности, при снижении напряжения кислорода в кро-ви. Молекула 2, 3-дифос-фоглицерата способна внедряться в центральную часть молекулы гемоглобина, что приводит к снижению сродства гемоглобина к кислороду. Кривая диссоциации смещается вправо. Кислород легче переходит в ткани. 2. Сродство гемоглобина к кислороду снижается при увеличении концентрации Н4' и двуокиси углерода (рис. 152). Кривая диссоциации оксигемоглобина в этих условиях также смещается вправо. 3. Подобным же образом действует на диссоциацию оксигемоглобина повышение температуры. Нетрудно понять, что эти изменения сродства гемоглобина к кислороду имеют важное значение для обеспечения снабжения им тканей. В тканях, в которых процессы обмена веществ протекают интенсивно, концентрация двуокиси углерода и кислых продуктов увеличивается, а температура повышается. Это ведет к усилению диссоциации оксигемоглобина.
Гемоглобин крови плода (HbF) обладает значительно большим сродством к кислороду, чем гемоглобин взрослых (НЬА). Кривая диссоциации HbF по отношению к кривой диссоциации НЬА сдвинута влево.
В волокнах скелетных мышц содержится близкий к гемоглобину миоглобин. Он обладает очень высоким сродством к кислороду.
Количество кислорода в крови. Максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. Для ее определения кровь насыщают кислородом воздуха. Кислородная емкость крови зависит от содержания в ней гемоглобина.
|
Один моль кислорода занимает объем 22,4 л. Грамм-молекула гемоглобина способна присоединить 22 400Х4== 89 600 мл кислорода (4 — число гемов в молекуле гемоглобина). Молекулярная масса гемоглобина — 66 800. Значит, 1 г гемоглобина способен присоединить 89 600:66 800== 1,34 мл кислорода. При содержании в крови 140 г/л гемоглобина кислородная емкость крови будет 1,34-. 140=== 187,6 мл, или около 19 об. % (без учета небольшого количества физически растворенного в плазме кислорода).
В артериальной крови содержание кислорода лишь немного (на 3—4%) ниже кислородной емкости крови. В норме в 1 л артериальной крови содержится 180—200 мл кислорода. При дыхании чистым кислородом его количество в артериальной крови практически соответствует кислородной емкости. По сравнению с дыханием атмосферным воздухом количество переносимого кислорода увеличивается мало (на 3—4%), но при этом возрастают напряжение растворенного кислорода и способность его диффундировать в ткани.
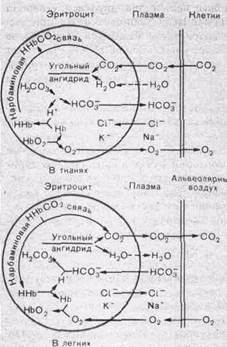
| Рис.153. Процессы, происходящие в эритроците при поглощении или отдаче кровью кислорода и двуокиси углерода (схема). |
Венозная кровь в состоянии покоя содержит около 120 мл/л кислорода. Таким образом, протекая по тканевым капиллярам, кровь отдает не весь кислород. Часть кислорода, поглощаемая тканями из артериальной крови, называется коэффициентом утилизации кислорода. Для его вычисления делят разность содержания кислорода в артериальной и венозной крови на содержание кислорода в артериальной крови и умножают на 100. Например: (200— 120) :200-100=40%. В покое коэффициент утилизации кислорода колеблется от 30 до 40%. При тяжелой мышечной работе он повышается до 50— 60%.
Дата добавления: 2016-03-27; просмотров: 1753;