Образование субстратов, необходимых для синтеза ЖК
Образование и транспорт Ацетил-КоА. В реакциях гликолиза из глюкозы образуется ПВК, который поступает в матрикс митохондрий и превращается в Ацетил-КоА с участием ПВК ДГ. Так как внутренняя мембрана митохондрий непроницаема для Ацетил-КоА, поэтому он при участии цитратсинтазы конденсируется с ЩУК с образованием цитрата:
Ацетил-КоА + Оксалоацетат → Цитрат + HS-КоА.
Затем транслоказа переносит цитрат в цитоплазму. Перенос цитрата в цитоплазму происходит только при увеличении количества цитрата в митохондриях, когда изоцитратдегидрогеназа и α-кетоглутаратдегидрогеназа ингибированы высокими концентрациями НАДН2 и АТФ (при избытке углеводов и низком энергопотреблении).
В цитоплазме цитрат расщепляется под действием фермента цитрат-лиазы:
Цитрат + HSKoA + АТФ → Ацетил-КоА + АДФ+ Pн + ЩУК
Образование НАДФН2.
1) ЩУК в цитоплазме превращается в малат под действием малат ДГ, малат под действием малик-фермента превращается в ПВК, при этом образуется НАДФН2. ПВК транспортируется обратно в матрикс митохондрий;
2) НАДФН2 образуется в цитоплазме из глюкозы в окислительных реакциях ПФШ;
3) Цитрат изомеризуется в изоцитрат, который дегидрируется цитозольной НАДФ-зависимой ДГ до α-КГ с образованием НАДФН2. α-КГ переноситься в матрикс митохондрий.
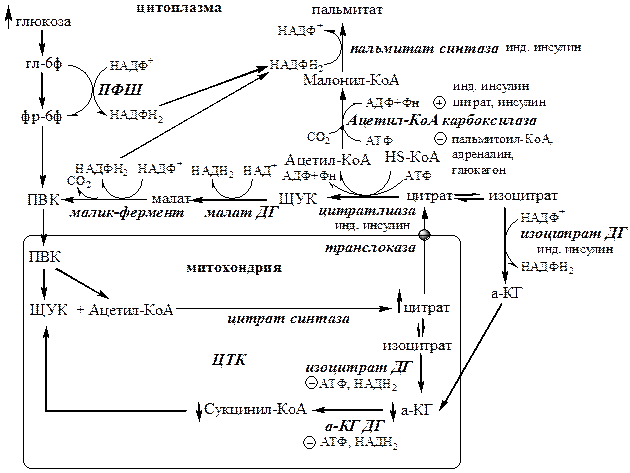 Синтез пальмитиновой кислоты
Синтез пальмитиновой кислоты
Образование малонил-КоА
Первая реакция синтеза ЖК — превращение ацетил-КоА в малонил-КоА. Это регуляторная реакция в синтезе ЖК катализируется ацетил-КоА-карбоксилазой.
Ацетил-КоА-карбоксилаза состоит из нескольких субъединиц, содержащих биотин.
Реакция протекает в 2 стадии:
1) СО2 + биотин + АТФ → биотин-СООН + АДФ + Фн
2) ацетил-КоА + биотин-СООН → малонил-КоА + биотин
Ацетил-КоА-карбоксилаза регулируется несколькими способами:
1) Ассоциация/диссоциация комплексов субъединиц фермента. В неактивной форме ацетил-КоА-карбоксилаза представляет собой комплексы, состоящих из 4 субъединиц. Цитрат стимулирует объединение комплексов, в результате чего активность фермента увеличивается. Пальмитоил-КоА вызывает диссоциацию комплексов и снижение активности фермента;
2) Фосфорилирование/дефосфорилирование ацетил-КоА-карбоксилазы. Глюкагон или адреналин через аденилатциклазную систему стимулируют фосфорилирование субъединиц ацетил-КоА карбоксилазы, что приводит к ее инактивации. Инсулин активирует фосфопротеинфосфатазу, ацетил-КоА карбоксилаза дефосфорилируется. Затем под действием цитрата происходит полимеризация протомеров фермента, и он становится активным;
3) Длительное потребление богатой углеводами и бедной липидами пищи приводит к увеличению секреции инсулина, который индукцирует синтез ацетил-КоА-карбоксилазы, пальмитатсинтазы, цитратлиазы, изоцитратдегидрогеназы и ускоряет синтез ЖК и ТГ. Голодание или богатая жирами пища приводит к снижению синтеза ферментов и, соответственно, ЖК и ТГ.
Образование пальмитиновой кислоты
После образования малонил-КоА синтез пальмитиновой кислоты продолжается на мультиферментном комплексе — синтазе жирных кислот (пальмитоилсинтетазе).
Пальмитоилсинтаза - это димер, состоящий из двух идентичных полипептидных цепей. Каждая цепь имеет 7 активных центров и ацилпереносящий белок (АПБ). В каждой цепи есть 2 SH-гpyппы: одна SH-гpyппa принадлежит цистеину, другая — остатку фосфопантетеиновой кислоты. SH-группа цистеина одного мономера расположена рядом с SH-группой 4-фосфопантетеината другого протомера. Таким образом, протомеры фермента расположены «голова к хвосту». Хотя каждый мономер содержит все каталитические центры, функционально активен комплекс из 2 протомеров. Поэтому реально синтезируются одновременно 2 ЖК.
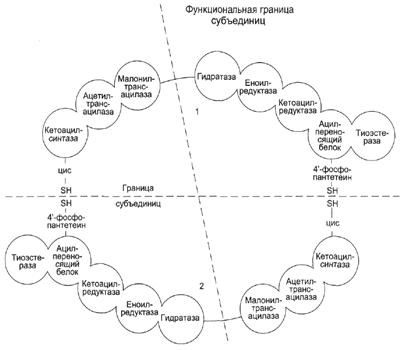
Этот комплекс последовательно удлиняет радикал ЖК на 2 атома С, донором которых служит малонил-КоА.
Реакции синтеза пальмитиновой кислоты
1) Перенос ацетила с КоА на SH-группу цистеина ацетилтрансацилазным центром;
2) Перенос малонила с КоА на SH-группу АПБ малонилтрансацилазным центром;
3) Кетоацилсинтазным центром ацетильная группа конденсируется с малонильной с образованием кетоацила и выделением СО2.
4) Кетоацил восстанавливается кетоацил-редуктазой до оксиацила;
5) Оксиацил дегидратируется гидратазой в еноил;
6) Еноил восстанавливается еноилредуктазой до ацила.
В результате первого цикла реакций образуется ацил с 4 атомами С (бутирил). Далее бутирил переносится из позиции 2 в позицию 1 (где находился ацетил в начале первого цикла реакций). Затем бутирил подвергается тем же превращениям и удлиняется на 2 атома С (от малонил-КоА).
Аналогичные циклы реакций повторяются до тех пор, пока не образуется радикал пальмитиновой кислоты, который под действием тиоэстеразного центра гидролитически отделяется от ферментного комплекса, превращаясь в свободную пальмитиновую кислоту.
Суммарное уравнение синтеза пальмитиновой кислоты из ацетил-КоА и малонил-КоА имеет следующий вид:
CH3-CO-SKoA + 7 HOOC-CH2-CO-SKoA + 14 НАДФН2 → C15H31COOH + 7 СО2 + 6
Н2О + 8 HSKoA + 14 НАДФ+
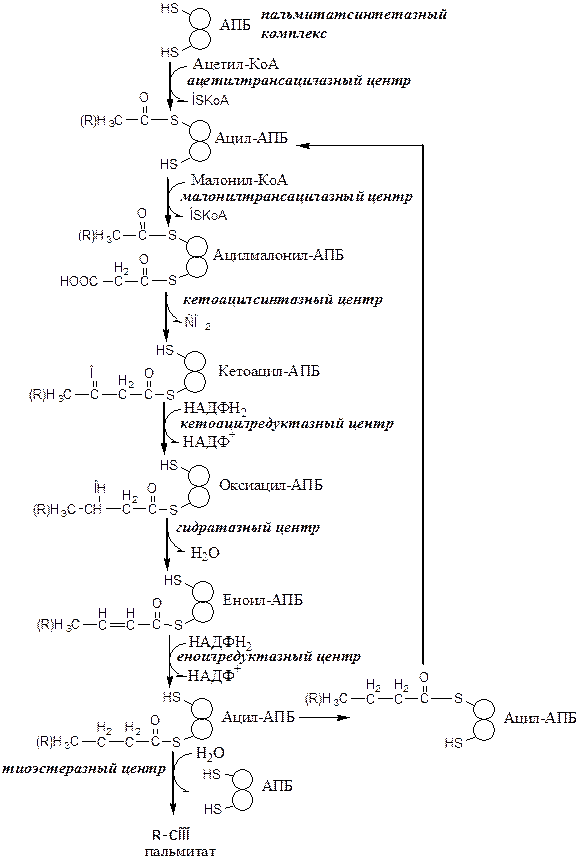
Дата добавления: 2016-02-13; просмотров: 795;