Уровень недифференцированной чувствительности
Самый низший, зародышевый уровень развития психического, отвечающий, согласно А.Н. Леонтьеву, недифференцированной чувствительности, следует непосредственно за допсихической формой жизни и допсихической формой отражения. Соответственно этому фаза фоновой активности (всегда отвечающая, как было показано в § 3 главы 1, предыдущему уровню развития) осуществляется здесь на допсихическом уровне и связана с физиолого-биохимическими факторами, т.е. может быть названа поисковой лишь условно. К.Э.Фабри отмечает, что на «исходном уровне развития психического отражения первичные формы внешней двигательной активности... детерминировались только раздражимостью».
Можно полагать, что поисковая активность побуждалась на данном уровне прекращением ассимилятивных биохимических реакций вследствие происходившего по тем или иным причинам отделения живого организма от содержащих жизненно необходимые свойства или компоненты частей среды. В простейшем случае (при незначительном пространственном разъединении) она, по-видимому, выражалась в беспорядочных хаотичных движениях, в конце концов приводивших к требуемому для собственно активной фазы деятельности сближению с предметом потребности. Эти движения, по всей вероятности, имели либо биохимическую, либо физико-химическую природу.
Очевидно, что первичная недифференцированная чувствительность направлена на необходимые для поддержания жизни свойства среды, приобретающие сигнальный характер. Собственно активная фаза деятельности проявляется в реакции на эти свойства, наступающей при определенном (на первых этапах развития, по-видимому, очень тесном) сближении с предметом потребности. По его достижении все как бы возвращается на круги своя, реакция (R) фактически знаменует восстановление внутренней деятельности. Внешняя деятельность является здесь как бы простым придатком внутренней (ассимилятивных процессов), составляющей на уровне недифференцированной чувствительности основную, существенно преобладающую часть жизнедеятельности.
Итак, общую структуру жизнедеятельности на уровне недифференцированной чувствительности можно представить в следующем виде: актуализация потребности (прекращение ассимилятивных процессов) — фоновая активность - стимул-реакция (S-R) — достижение предмета потребности (восстановление ассимилятивных процессов). Стимул — реакция (S-R) выступают как прообраз рефлекторного акта («прарефлекс»), который состоит из непосредственно связанных афферентного и эффекторного звеньев. Отметим, что собственно активная фаза внешней деятельности осуществляется лишь на границе с внутренней, в «прарефлексе».
Психическое отражение на данном уровне выступает как чувствительность к жизненно значимым свойствам среды. Она является и внутренней стороной результата отражения. В то же время уже здесь можно выделить и внешнюю сторону результата отражения — свойства среды, имеющие сигнальное значение.
Среди современных живых организмов не встречаются животные, отвечающие уровню недифференцированной чувствительности в его первоначальном виде. К.Э.Фабри указывал, что «строение и поведение современных «простейших» являются уже намного более сложными, чем у их ископаемых предков. Он полагает, что при зарождении животной жизни существовал более низкий, чем у современных одноклеточных, действительно первоначальный уровень развития.
В то же время многие современные животные близки к уровню недифференцированной чувствительности. Это простейшие (амебы, инфузории, жгутиковые и т.д.), многие низшие многоклеточные (губки, коралловые полипы и др.), для которых К.Э. Фабри выделил низший уровень стадии элементарной сенсорной психики.
Выше отмечалось, что на первоначальном уровне психической (животной) жизни фаза фоновой активности еще не связана с психикой и остается обусловленной допсихическими формами отражения. Поведение современных простейших подтверждает указанный вывод.
Способы передвижения простейших весьма разнообразны. Некоторые пассивно перемещаются вместе с толщей воды, в которой они находятся, есть простейшие, перемещающиеся реактивным способом. Амебы двигаются посредством перераспределения плотности протоплазмы, как бы «переливая» ее из одного участка тела в другой. Большинство же простейших передвигается с помощью жгутиков или ресничек, представляющих собой волосовидные плазматические выросты. Однако, как почеркивал К.Э. Фабри, независимо от перечисленных способов, движения простейших (кинезы) в основном определяются «непосредственным воздействием градиентов интенсивности биологически значимых внешних факторов», таких, как химический, температурный и т.п. С какими-либо психическими процессами локомоция (передвижение) простейших в фазе фоновой активности не связана.
Для простейших характерны два вида кинезов — ортокинез и клинокинез. При ортокинезе животные осуществляют поступательное движение с переменной скоростью, определяемой градиентом воздействующего внешнего фактора. При клинокинезе изменяется направление движения. Изменение направления движения совершается хаотично, так что животное «находит» наиболее благоприятный вектор движения после «проб и ошибок».
Кинезы носят ненаправленный характер и не обеспечивают подлинной ориентации. Ориентирующие процессы, вносящие адекватную коррекцию в хаотическое изменение характера движения, называются таксисами.
Таксисы простейших, как и их кинезы в целом, не связаны с проявлением психического. Показательна в этом отношении тенденция к движению вверх инфузории туфельки, не имеющей гравитационной чувствительности. Ориентация движения туфельки определяется действием содержимого пищеварительных вакуолей, которое давит на их нижнюю часть. Если над туфелькой, проглотившей порошок железа, поместить магнит, она начинает двигаться не вверх, а вниз: порошок давит не на нижние, а на верхние части вакуолей, что приводит к обратной ориентации движения.
Не связаны с проявлением психики и реакции простейших на температуру. К.Э. Фабри отмечает, что «здесь непосредственно необходимая для поддержания жизни энергия еще тождественна опосредствующей энергии, сигнализирующей о наличии жизненно необходимого компонента среды». Какие-либо специфические терморецепторы у простейших отсутствуют. К.Э. Фабри полагает, что реакции простейших на температуру сходны с биохимическими реакциями типа ферментативных процессов.
Собственно активная фаза жизнедеятельности простейших, связанная с зачаточными проявлениями психического, осуществляется в непосредственной близости от предмета потребности. По имеющимся данным, амеба реагирует на пищу с расстояния 20—30 мкм. Активно реагировать на предмет пищевой потребности способны и хищные инфузории. К собственно активной фазе деятельности простейших относятся и фобические реакции, проявляющиеся, например, у амеб и инфузорий. Так, при контакте с твердым предметом либо оказавшись в каких-то других неблагоприятных условиях туфелька прекращает движение и «съеживается», что обычно предшествует выделению стрекательных капсул.
Пластичность поведения — способность к индивидуальному изменению врожденного видотипичного поведения у простейших выражена весьма слабо. Она ограничивается наиболее примитивной формой — привыканием. В опытах немецкого ученого Ф. Брамштедта парамеции определенное время держались в сосуде треугольного или квадратного сечения, а затем перемещались в более крупный сосуд круглого сечения. В нем они продолжали сохранять привычное движение по траектории, повторяющей периметр прежнего сосуда (рис. 1.3).
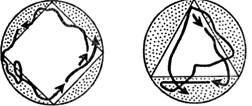
Рис. 1.3. Привыкание у инфузории туфельки (по Ф. Брамштедту)
Сенсорный уровень
Сенсорный уровень развития психики связан с преобразованием первоначально недифференцированной чувствительности в дифференцированные ощущения. Как показал А.Н. Леонтьев, последние возникают при освоении животными организмами все более сложных и многообразных сред обитания.
Среди других психических процессов ощущения выделяются своей парадоксальной специфичностью. Л.М. Веккер отмечает, что, с одной стороны, ощущение формируется в пространстве нейрофизиологического субстрата и представляет собой упорядоченное множество состояний последнего, а с другой — дает «представление о свойствах объектов именно как собственных свойствах внеположенных по отношению к субъекту предметов».
Таким образом, характерной чертой ощущения является его двойственность, принадлежность одновременно и к физиологическим, и к психическим явлениям. В то же время уже в рамках психических процессов ощущения выступают как явления, по существу примыкающие к эмоциям. Не случайно понятия «ощущать» и «чувствовать» нередко взаимозаменяемы: способность ощущать называется чувствительностью, органы ощущений — органами чувств. Можно сказать о чувстве горечи во рту и об ощущении радости и т.д. Известно понятие «эмоционального тона ощущений» (В. Вундт, П.В. Симонов, В.К. Вилюнас). Все это позволяет говорить о размытости границы между ощущениями и эмоциональными явлениями.
Ощущение можно рассматривать как психофизиологический процесс, состоящий из двух начал: 1) «отпечатка» раздражителя, «упорядоченного множества состояний» нейрофизиологического субстрата и 2) эмоционального эквивалента, выступающего в виде «чувственного представления» (В. Вундт).
Независимо от того, является ли то или иное ощущение контактным, дистантным или обонятельным, оно, с одной стороны, — отпечаток, носитель непосредственной материальной связи субъекта с объектом; с другой стороны, как подчеркивает Л.М. Веккер, оно обладает «объектной специфичностью», которую вслед за В.К. Вилюнасом можно связать с вундтовскими слияниями «простых чувств». В то же время эмоциональное начало ощущения может рассматриваться не как его эмоциональный тон, а в качестве непосредственной составляющей. Согласно распространенным представлениям (У. Джемс, У. Мак-Дауголл, В.К. Вилюнас) в рефлекторных актах инстинктивного поведения между стимулом и реакцией возникает эмоция, которая и служит непосредственным побуждением к действию. Данную трактовку можно с полным основанием распространить и на ощущения, которые, как известно, являются целостными рефлекторными актами. В этом случае вундтовские «чувственные представления» отдельных специфических качеств раздражителей, появляющиеся при ощущениях, могут рассматриваться как первичные элементарные эмоции, возникающие в результате синтезирующего процесса в центральных и эфферентных звеньях соответствующих рефлексов.
Согласно В. Вундту, «простые чувства» образуют одно целое связное многообразие», которое может быть выражено в трех измерениях — возбуждение, напряжение и удовольствие, каждое из которых имеет как положительный, так и отрицательный вектор. В. Вундт приводит доводы, показывающие «непосредственное родство» чувственных тонов различных ощущений: «Нам кажется, что низкие тона соответствуют темным цветам и черному, высокие тона — светлым цветам и белому. Резкий звук, например звук трубы, и цвета из возбуждающего ряда — желтый и светло-красный -- соответствуют друг другу... Подобные же сравнения... делаем мы и тогда, когда различаем теплый и холодный цвета, или говорим: «резкий звук» и «насыщенный цвет» и т.д.».
Несомненная аналогия, даже взаимозаменяемость звуковых и цветовых ощущений хорошо показана В.Г. Короленко в повести «Слепой музыкант». К этому можно добавить такую же аналогию вкусовых и обонятельных, зрительных и осязательных ощущений и т.д. Как известно, слепоглухие от рождения люди тем не менее обладают целостной картиной мира, основанной на имеющихся у них других ощущениях. Все это свидетельствует о взаимозаменяемости ощущений, что обусловлено наличием в них наряду с физиологическим эмоционального начала.
Общая структура жизнедеятельности на сенсорном уровне может быть представлена следующим образом:
Актуализация потребности — фоновая (поисковая) активность — S-E-R — достижение предмета потребности.
S-E-R (стимул — эмоция — реакция) представляет собой рефлекторный акт ощущения с соответствующими эффекторными звеньями. Связь между потребностью и ее предметом усложняется, теперь она включает фоновую активность и целостный рефлекс, развившийся из простейшего «прарефлекса» предыдущего уровня. Меняется и характер фоновой активности. На данном уровне она определяется общей чувствительностью к тем свойствам среды, которые прежде обусловливали собственно активность. Прежняя собственно активная фаза становится фоновой активностью, ее составляющей. Центральной собственно активной фазой становится дифференцированное ощущение с дополнительными эффекторными звеньями.
Отражательный аспект связи субъекта с объектом имеет двойственный характер: с одной стороны, непосредственное взаимодействие с объектом (раздражители — рецепторы), а с другой -- психическое (эмоциональное) начало ощущения, появляющееся вследствие синтеза в центральном и эфферентном звеньях рефлекса. Результат отражения соответственно предстает, с одной стороны, как непосредственный «отпечаток» жизненно значимых свойств объекта в нейрофизиологическом субстрате (внутренний аспект), а с другой — как «чувственное представление» этих свойств (внешний аспект).
Отметим, что на сенсорном уровне связь субъекта с объектом, оставаясь непосредственной, значительно отличается от непосредственного взаимодействия со средой, характерного для предыдущего уровня. Эта связь в несравненно большей степени организуется психическим началом. Уже на уровне недифференцированной чувствительности можно констатировать обусловленность отражения биологической «смысловой связью» (А.Н. Леонтьев), в результате образования которой «соответствующая потребность... как бы «узнает себя» в данном предмете-раздражителе, конкретизируется в нем». На сенсорном уровне эта связь функционально оформляется как целостный рефлекторный процесс, который можно рассматривать в качестве основного общеорганизующего начала жизнедеятельности данного этапа филогенеза.
Между уровнем недифференцированной чувствительности и сенсорным уровнем — уровнем дифференцированных ощущений — лежит долгий путь эволюции. Отражением этого пути являются многие современные животные, находящиеся на промежуточных уровнях развития. К ним относятся большинство кишечнополостных (в том числе гидры, медузы, некоторые актинии), низшие черви и ряд других беспозвоночных. Следует иметь в виду, что все ныне живущие животные в филогенетическом отношении представляют собой боковые ветви эволюционного развития, далеко не всегда совпадающие с выделенными уровнями психического отражения (рис. 1.4).
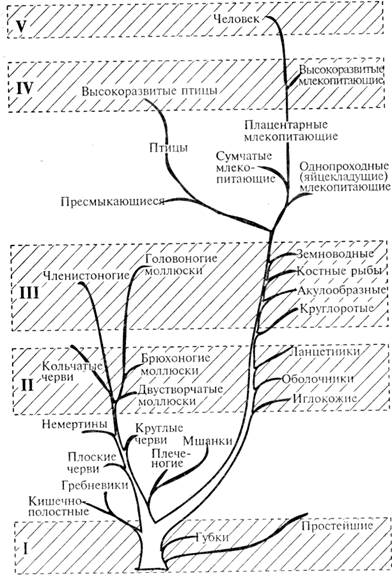
Рис. 1.4. Современные животные и уровни психического развития в филогенезе
Тем не менее большая группа низших беспозвоночных — высшие (кольчатые) черви, иглокожие, двустворчатые моллюски, брюхоногие моллюски (улитки) и многие другие в целом соответствуют сенсорному уровню психического. В наибольшей степени это относится к большинству видов кольчатых червей.
Двигательная активность кольчатых червей достаточно многообразна и сложна. Локомоция обеспечивается хорошо развитой мускулатурой, состоящей из так называемого кожно-мышечного мешка. Его внешний слой представлен кольцевыми волокнами, внутренний — продольными. При передвижении червь ритмично сокращает продольную и кольцевую мускулатуру, в результате чего попеременно вытягиваются и сокращаются отдельные сегменты.
У кольчатых червей впервые в эволюции появляются парные конечности — параподии. На каждом сегменте имеются парные выросты, обычно снабженные опорными щетинками. Специальные мышцы могут двигать их вперед или назад. Параподии также снабжены органами тактильной и химической чувствительности, имеющими вид щупалец. На головном конце параподии служат ротовыми органами. У многих кольчатых червей, в том числе у дождевого, параподии редуцированы до невидимых невооруженным глазом пучков щетинок, парно расположенных на каждом сегменте. Тем не менее в сочетании с сокращениями кожно-мышечного мешка они способствуют передвижению.
Органы чувствительности типичных представителей сенсорного уровня психики как морфологически, так и функционально дифференцированы слабо. К.Э. Фабри полагает, что первоначально они представляли собой рецепторные клетки с повышенной общей (нерасчлененной) чувствительностью. Согласно принятым представлениям, все органы чувств в филогенезе возникли на основе органов осязания. У наиболее примитивных животных способностью осязания обладают все клетки поверхности тела. Затем (уже у кишечнополостных) появляются скопления специализированных осязательных клеток — первичные органы осязания. Нередко они одновременно выполняют и обонятельную функцию. Морфологическая и функциональная слитность органов осязания и обоняния является характерной особенностью большинства представителей сенсорного уровня развития психики.
В то же время основная жизнедеятельность (добывание пиши) животных данного уровня осуществляется с использованием только одного из органов чувств — осязания, реже обоняния или зрения. Органы зрения у этих животных либо отсутствуют вовсе, либо представляют собой лишь органы светочувствительности, позволяющие отличать свет от тьмы и определять расположение источника света.
Жизнедеятельность большинства кольчатых червей состоит главным образом из врожденных стереотипных программ (заглатывание субстрата, содержащего пищевые компоненты, процеживание воды с мелкими животными и т.п.). Ее можно представить как постоянное повторение одного и того же рефлекторного акта ощущения (обычно осязательного), дополненного соответствующими эффек-торными звеньями (заглатывание, процеживание и т.д.). Легко видеть, что эта монотонная рефлекторная деятельность, практически лишенная ситуативности, является собственно активной фазой жизнедеятельности, отвечающей психическому отражению максимально возможного для данных животных уровня.
Фаза фоновой активности связана у большинства кольчатых червей с поиском субстрата, содержащего пищевые компоненты. В общем случае она не предполагает участия дифференцированных ощущений и, по всей вероятности, обычно осуществляется на основе недифференцированной или слабо дифференцированной чувствительности.
Отметим, что на данном уровне развития психики основная часть жизнедеятельности представлена фазой собственно активности, состоящей из однообразных рефлекторных актов. Необходимость в поисковой активности возникает эпизодически, а в ряде случаев (например, у кольчатых червей, проводящих весь свой жизненный цикл в иле, у дождевых червей в богатых перегноем почвах и т.д.) вообще может не возникать.
Пластичность поведения у животных сенсорного уровня развития психики выражена слабо. Как и для простейших, для них характерна самая примитивная форма пластичности поведения — привыкание. Врожденные реакции на те или иные раздражители, остающиеся без последствий, после многократных повторных воздействий прекращаются. Так, дождевой червь, реагирующий на внезапное уменьшение интенсивности освещения движениями бегства, через определенное количество повторений прекращает эти реакции.
Тем не менее уже на данном уровне появляются зачатки способности к ассоциативным связям, хотя последние формируются с трудом и в ограниченных пределах. В опытах Р. Иеркса дождевые черви помещались в Т-образный лабиринт, где в одном из двух возможных «ходов» получали электрический удар (рис. 1.5). Через 120—180 повторений они начинали постоянно выбирать безопасное направление. Улитки в аналогичных опытах приучались выбирать нужное направление через 60 повторений, запоминая правильное решение на 30 дней.
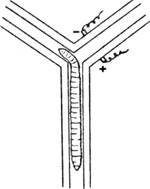
Рис. 1.5. Научение у дождевого червя (по К.Э. Фабри)
В экспериментах с червями было установлено также, что они способны научиться правильно ориентироваться в лабиринте даже без предварительно удаленных передних сегментов. Таким образом, каждый из сегментов в значительной степени автономно обеспечивает жизнедеятельность животных. Это свидетельствует о еще сравнительно малом влиянии начавшегося процесса цефализации (обособления головной части) на поведенческие акты. Тем не менее определенная роль передних сегментов все же несомненна: после разрезания дождевого червя пополам передняя половина зарывается в землю, совершая такие же координированные движения, как и целый червь, задняя же часть беспорядочно извивается.
Дата добавления: 2016-03-04; просмотров: 1052;