Как получить новое, унаследовав старое? 3 страница
Примерно таким же образом, судя по всему, шел и процесс возникновения человека с его видоспецифической коммуникативной системой – языком.
С наступлением эпохи более холодного и сухого климата перед приматами открывается новая возможность – огромные (и все увеличивающиеся) пространства, занимаемые полуоткрытыми (редколесье) и открытыми (саванна) ландшафтами. До миоцена (а именно в конце миоцена появляются первые представители клады человека), судя по палеонтологическим данным, ничего похожего на степи или саванны в Африке не было99 – саванны возникают в процессе совместной эволюции “злаков и крупных травоядных млекопитающих”100 . Их начинают осваивать самые разные группы животных – таким образом в интервале 3–2,5 млн. лет назад появляются десятки новых видов грызунов и крупных копытных101 . Для привычных к условиям влажного тропического леса приматов такие местообитания представляются скорее неудобными, поскольку там хуже, чем в лесу, работают привычные стратегии добывания пищи, поиска укрытий и т. п. Но по количеству кормовых ресурсов эти места не настолько безнадежны, чтобы особи, не сумевшие встроиться в лесные группировки, немедленно погибали. Пищи в редколесье и даже в саванне вполне достаточно – надо только суметь ею воспользоваться. Но условия ее поиска в такого рода мозаичных ландшафтах совершенно не таковы, как во влажных тропических ландшафтах, где обитали предки гоминид102 . Источники пищи существенно меняются в разные сезоны – во влажное время года в изобилии имеется растительная пища, термиты, птичьи яйца и т. д., в сухой сезон можно есть семена и корнитрав, ловить рыбу в высыхающих водоемах, питаться мясом – либо охотясь на травоядных, сосредотачивающихся в это время года около источников воды, либо поедая трупы павших животных104 . Длина влажного и сухого сезонов в разные годы неодинакова, что делает жизнь в саванне еще более разнообразной и непредсказуемой105 . Кроме того, в саваннах, где было много травянистой растительности и, соответственно, много травоядных животных, “увеличилось видовое разнообразие и общая численность хищников. Такое положение вещей с неизбежностью приводило к ситуации, основным плюсом которой было появление новых источников пищи, а основным недостатком – повышение риска встречи с хищником”106 , и это тоже было очень существенным фактором, к которому гоминиды должны были адаптироваться.

Рис. 5.4. Модель оптимизации и приспособления к условиям для пищевого поиска гоминид на протяжении сезонного цикла (с учетом сезонных изменений доступности пищевых ресурсов), предложенная Р. Фоули 103 .
Вообще, открытые ландшафты, по‑видимому, дают больше, чем лесные, разнообразных условий среды, к которым можно приспосабливаться. Если, например, рассмотреть эволюцию в семействе полевок107 , то можно увидеть, что род рыжих полевок, живущих в лесу, содержит сравнительно немного видов, и все они довольно архаичны, то есть сформировались достаточно давно и с тех пор не менялись. Напротив, серые полевки, освоившие луговые местообитания, демонстрируют целый спектр сравнительно молодых видов – это значит, что им приходилось то и дело приспосабливаться к меняющимся условиям среды.
О том, что экологическая ниша, которую начали осваивать гоминиды, была для них новой, свидетельствует, в частности, полиморфизм ранних представителей клады человека (независимо от того, являются ли найденные останки действительно принадлежащими столь многим разным видам или даже родам или в ряде случаев демонстрируют лишь разнообразие локальных вариаций в рамках одного рода или вида). Возникновение любого нового таксона “до некоторой степени аналогично проникновению на мало заселенные острова”108 (т. е. в новую экологическую нишу), и на начальных этапах его существования имеет место “бурная дивергентная эволюция”109 , причем ранние члены таксона даже на низких таксономических уровнях резко различаются между собой, “порой по признакам, которые позже будут характеризовать таксоны гораздо более высокого ранга”110 (так называемый “закон архаического многообразия”). Движение от разнообразия к единообразию в процессе происхождения человека иллюстрируют в том числе и следующие факты. У Homo habilis наблюдается значительная вариативность по объему мозга (от 500 до 800 см3) – и на следующую ступень эволюции “переходят” лишь обладатели самого большого мозга (в итоге у архантропов он достигает более 900 см3). Точно так же у Homo heidelbergensis сильно варьировали возможности слухового анализатора (см. рис. 3.6 в гл. 3) – и для перехода на следующую ступень эволюции были отобраны обладатели лишь одного из возможных вариантов. Видимо, отбор шел на приспособленность к определенным поведенческим свойствам, а сами свойства появлялись в ответ на некий эволюционный вызов. Примерно так же происходило образование нового вида тлей в опытах Георгия Христофоровича Шапошникова111 : при насильственной пересадке в другую экологическую нишу (на другое растение) наблюдалось сначала повышение вариативности, а потом (за 170 поколений) из этого множества вариантов отобрались наиболее приспособленные, и появился новый вид (не только отличающийся по своему строению, но и не скрещивающийся с видом‑предком).
О том, какую именно нишу стали осваивать далекие предки человека, существуют разные мнения. Одни исследователи полагают, что гоминиды занимались поеданием падали – в это время сильно уменьшилось число видов крупных гиен112 , так что невостребованных трупов должно было быть в достатке. Кроме того, гоминиды за счет того, что отсутствие волос, обилие потовых желез и выпрямленная походка предохраняли их от перегрева, могли быть активны в жаркие дневные часы, избегая тем самым конкуренции с другими любителями мяса (и хищниками, и падальщиками), которые в это время отдыхают113 . По мнению других, они могли охотиться. Но в любом случае, “наблюдаемые увеличения общих размеров тела, объема мозга, продолжительности жизни не могли иметь места без смены пищевой стратегии, а смена пищевой стратегии означает переход в новую экологическую нишу”114 .
Для такого перехода необходимо развитие нужных специализаций. Но природа, как уже говорилось, не производит неограниченного спектра форм, из которых естественный отбор мог бы выбирать наилучшие – эволюция обычно идет по пути развития того, что у организмов уже существует. Для нее оказывается существенно проще все более и более усиливать уже существующую тенденцию к той или иной специализации, чем отказываться от нее и переходить к специализации совершенно иного типа, поэтому, например, среди кошачьих появляются все более и более эффективные хищники, но не возникает травоядных форм. Эволюция приматов не является исключением из общего правила.
Как отмечают Бутовская и Файнберг, предковые для го‑минид формы скорее всего были всеядны и не имели выраженных специализаций к какому‑либо определенному типу добычи пищи115 . У них “не было ни быстроты стайных хищников, ни их зубов, ни когтей для охоты”116 , ни таких развитых анатомических приспособлений для питания грубой растительной пищей, как у копытных. Конкурировать с другими представителями фауны наши предки могли прежде всего за счет способности быстро адаптироваться к окружающей среде посредством поведения. Поведенческое приспособление имеет то преимущество перед приспособлением анатомо‑физиологическим, что осуществляется в более короткие сроки, позволяет более оперативно реагировать на изменения окружающей среды, быстро осваивать новые условия (и тем самым расширять свою экологическую нишу).

Рис. 5.5. Чтобы научиться лакомиться сливками, никакие мутации не нужны. Как показали Дэвид Шерри и Беннет Галеф 125 , не нужно даже наблюдать, как проклевывают крышку: одного вида бодро скачущего поблизости сородича достаточно, чтобы побудить синицу продырявить крышку и добраться до сливок.
В нынешнее время мы можем наблюдать это на примере освоения городов различными видами птиц117 . Например, грач (Corvus frugilegus ) освоил города Приднестровья за 15 лет118 , вяхирь (Columba palumbus ) за 10 лет смог сформировать жизнеспособную популяцию во Львове119 . Поведение городских птиц сильно отличается от поведения их диких сородичей: грачи зимой кормятся на тротуарах, почти под ногами у прохожих, подобно голубям и воробьям, ястребы‑тетеревятники (Accipiter gentilis ) осваивают охоту на чердаках, выслеживают добычу в густых сумерках и даже “мышкуют” среди травы120 . Подобные нетипичные для диких птиц варианты поведения распространяются за время гораздо меньшее, чем понадобилось бы в случае, если бы эти поведенческие характеристики возникали в результате мутаций. Как отмечает орнитолог Вадим Вадимович Корбут, “формирование урбанизированных популяций серой вороны и кряквы в Москве в 60–80‑е годы привело к нарастанию их численности на 2–3 порядка при жизни особей‑основателей”121 . Впрочем, следует отметить, что эти элементы поведения именно нетипичны, а не вовсе невозможны для соответствующих видов – было показано, что они входят в видовую норму реакции122 . Подобного же рода быстрое распространение поведенческих признаков наблюдали в сороковые годы в Англии на примере синиц‑лазоревок (Parus caeruleus ), за считанные годы научившихся открывать закупоренные фольгой молочные бутылки123 . Широко известны наблюдения за японскими макаками острова Кошима, самостоятельно додумавшимися до того, что клубни батата (сладкого картофеля) можно мыть в ручье124 . Сначала этот способ очистки пищи от грязи применяла только одна самка Имо {44} – изобретательница метода, потом постепенно он распространился по всей группировке.
Рис. 5.6. А потом Имо догадалась, что клубень станет вкуснее, если надкусить его и окунуть в соленую морскую воду. Другие макаки, конечно же, собезьянничали.
Отметим специально, что рассмотренные изменения в поведении – это не результат мутаций, возникших у единичной особи и затем распространившихся: для мутационного сценария необходимо существенно больше эволюционного времени.
Такого рода поведенческое приспособление может иметь важные эволюционные последствия: оно запускает механизм, известный как “эффект Болдуина” (по имени американского психолога Джеймса Марка Болдуина, сформулировавшего в 1896 году эту гипотезу). Выглядит он следующим образом. Пусть у каких‑то особей данного вида есть в арсенале некоторая модель поведения, которая приносит им выгоду – помогает более эффективно находить пищу, избегать опасностей и т. д. Это значит, что их генетические характеристики, как минимум, не препятствуют реализации этой модели. Обладатели этих генетических характеристик оставят больше потомства, значит, в следующем поколении особей, которые обладают способностью к этому поведению, будет больше, и снова максимум потомства оставят именно они. В наибольшем эволюционном выигрыше оказываются те, чьи гены не только не препятствуют полезному поведению, но прямо ему способствуют, причем вовсе не обязательно превращать это поведение в инстинкт – достаточно просто уметь (быть генетически предрасположенным) быстро и надежно ему обучаться127 . Подчеркнем специально, что, когда в регуляции развития особи достаточно большое участие принимает внешняя среда, генетически передается не само сложное поведение (в частности, коммуникативное), а предрасположенность к нему128 , к тем или иным реакциям индивида на явления окружающей действительности – т. е. в конечном счете к тому, чтобы некоторые связи между нейронами устанавливались быстрее и были более прочными, чем другие129 .
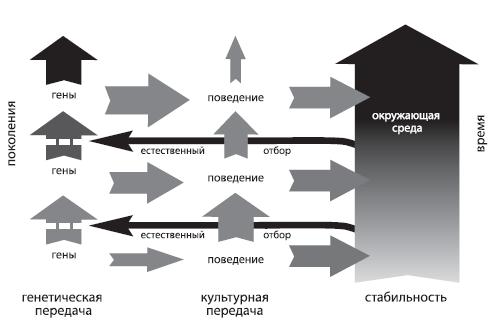
Рис. 5.7. Эффект Болдуина
Когда преимущества, которые дает то или иное поведение, достаточно серьезны, отбор благоприятствует тем популяциям, где особи, способные к нему, появляются чаще. Именно так, скорее всего, происходило и формирование языка – через постепенное накопление (сначала – в рамках видовой нормы реакции) тех, кто лучше умел доносить свои мысли до окружающих, и тех, кто был наиболее успешен в понимании. Если бы языковая способность возникла в результате мутации у единичной особи, язык не смог бы сформироваться – так же, как не складывался язык в никарагуанской школе для глухих, пока число стремящихся общаться между собой воспитанников не перевалило за две сотни131 .
Если то или иное поведение окажется полезным, будут закрепляться мутации, способствующие совместному наследованию всего того комплекса признаков, который обеспечивает данное поведение. Будут поддерживаться отбором и те мутации, которые будут изменять анатомию организма в сторону, благоприятствующую данному поведению. Например, успехи в ухаживательном поведении, включающем демонстрацию развернутого веером хвоста, формируют в конечном итоге хвост, весьма выигрышно выглядящий в развернутом виде. Вероятно, такого же рода эволюцию можно предполагать и для языка: если, например, стремление произносить звуковые сигналы максимально внятно и разборчиво приносило ощутимые выгоды в деле выживания и размножения, то отбором закреплялись такие анатомические свойства, которые максимально облегчали особям эту задачу (пусть даже в ущерб каким‑то другим жизненным интересам), – и в итоге люди получили свою опущенную гортань. Если выгодно было уметь передавать максимум информации за один раз, закрепились мутации, позволявшие (пусть с большей тратой ресурсов) более эффективно управлять дыханием, и т. д.
Этот механизм эволюции отличается от широко распространенных представлений, согласно которым сначала возникает мутация, а потом организмы пытаются извлечь из нее какую‑то пользу. В данном случае, наоборот, сначала формируются те или иные поведенческие навыки, и лишь потом те особенности организма, которые позволяют реализовывать их наилучшим образом, закрепляются генетически (что обеспечивает эффективное наследование поведения без прямого закрепления в генах конкретных поведенческих моделей). Согласно этому сценарию, мутации – необходимый компонент эволюционного процесса, но для каждого отдельного эволюционного события генетическое закрепление тех или иных характеристик фенотипа – не начальная, а, скорее, завершающая стадия. Какие именно фенотипические характеристики могут возникать в ходе эволюции подобным образом – вопрос сложный, и окончательного ответа на него в науке пока нет, но то, что в эволюции поведения эффект Болдуина играет значительную роль, по‑видимому, факт. А следовательно, для происхождения языка такой эволюционный механизм представляется более вероятным, нежели обратный (сначала – мутации, потом поведение).
Способности к поведенческому приспособлению имеются у многих животных, но человекообразные обезьяны продвинулись в этом значительно дальше других. Они способны не только осознать один элемент ситуации как средство обработки другого (например, ручей как средство для мытья клубней или камень как средство для того, чтобы расколоть орех), они – по крайней мере, шимпанзе – могут целенаправленно изготавливать орудия труда или переносить требующую обработки пищу в удобное для обработки место (см. гл. 3). Вообще, для видов, не имеющих выраженной специализации к определенному типу пищи, “характерны более разнообразный поведенческий репертуар и способность к полифункциональному применению элемента поведения, обобщению типичных для вида элементов в необычные комплексы‑комбинации”132 . Кроме того, всеядность имеет то преимущество перед растительноядностью, что позволяет иметь менее массивные кости челюстей и черепа в целом, а это, в свою очередь, позволяет увеличить объем мозга (а тем самым и возможность строить и хранить в памяти большее количество поведенческих программ) без создания дополнительных нагрузок на опорно‑двигательный аппарат, поскольку “при уменьшении объема кости на единицу объем мозга увеличивается на две единицы”133 (при сохранении той же массы).
Основная специализация приматов в природе – понимание причинно‑следственных связей, умение строить поведенческие программы, исходя не из каких‑то предзаданных шаблонов, а из представления о внутреннем устройстве наличной ситуации. Именно поэтому в условиях эксперимента они оказываются в состоянии догадаться, что высоко висящий банан можно достать, построив пирамиду из ящиков или соединив две палки, огонь спиртовки, мешающий добраться до лакомства, можно залить водой и т. д.
У людей способность понимать причинно‑следственные связи развита неизмеримо сильнее, чем у обезьян, следовательно, ее развитие занимало важное место в эволюции человека. В мозгу эту способность обеспечивают префронтальные формации коры. Так, обезьяна с поврежденной префронтальной корой не способна понять, что, если пищу на ее глазах переложили из ящика А в ящик В, то искать ее в ящике А бессмысленно, обезьяна же с неповрежденной корой легко справляется с подобными задачами134 . Человек (но не обезьяна) справляется и с задачей выбрать меньшее лакомство, если известно (из предыдущего опыта), что выбранное будет отдано другому (см. гл. 2). Иными словами, префронтальные формации коры обеспечивают возможность делать выводы из нескольких посылок, реагировать на стимул не непосредственно, а с учетом разных привходящих обстоятельств. Умения такого рода чрезвычайно важны для всеядного существа (например, они позволяют, увидев дерево с незрелыми плодами, вернуться к нему через некоторое время, когда плоды созреют). Они же играют значительную роль в обеспечении функционирования языка, позволяя понимать сообщения, содержащие несколько утверждений. Кроме того, префронтальные отделы коры помогают планировать сложное поведение (в том числе коммуникативное)135 , диктуют, что замечать, а что игнорировать136 , – это позволяет, в частности, опознавать одинаковые слова, произнесенные разными людьми, с разной громкостью, скоростью и т. п., как одно и то же слово. В гоминидной линии идет постоянное увеличение размеров (как абсолютных, так и относительных) префронтальной коры137 .
Согласно гипотезе американского антрополога Дуайта Рида138 , именно увеличение префронтальной коры явилось одним из важнейших факторов глоттогенеза. Дело в том, что в одном из ее участков (поля 9 и 46 по Бродману) расположен называемый “центральный исполнительный компонент” (англ. central executive component ) рабочей памяти, позволяющий удерживать внимание на нескольких объектах одновременно. Человек может оперировать одновременно примерно семью понятиями (точнее, 7±2), а шимпанзе – всего лишь двумя‑тремя. Как показывают наблюдения приматологов, шимпанзе сравнительно легко обучаются колоть орехи, манипулируя одновременно двумя предметами (орехом и камнем‑молотком), но с гораздо бóльшим трудом овладевают умением колоть орехи, требующим координированных действий с тремя предметами – орехом, камнем‑молотком и камнем‑наковальней139 . Увеличение объема рабочей памяти (англ. ST ‑WMC, short ‑term working memory capacity ) позволяет человеку, в отличие от обезьян, строить и понимать предложения, состоящие из многих слов, а также овладевать правилами их построения.
Главным эволюционным “козырем” гоминид был, пользуясь терминологией А.А. Зубова, “комплекс гоминизации”. В него входят “высокоразвитый мозг, анатомические особенности руки и возможность ее освобождения для использования, а затем изготовления орудий труда и охоты, стереоскопическое зрение, сложное групповое поведение, включая высокий уровень организации стада, достаточно развитые средства коммуникации, взаимодействие между индивидуумами (кооперацию), склонность к манипулированию предметами, гибкость поведения, высокий удельный вес поведенческих адаптаций в общей системе приспособительных реакций, повышенная роль прижизненного опыта в организации поведения и, соответственно, – возрастающая роль индивидуума в группе”140 . Как пишут Бутовская и Файнберг, “дифференциация поведения особей на индивидуальном уровне, связанная с уникальностью их прижизненного опыта в сочетании с индивидуальными психическими задатками… способствовала повышению пластичности функционирования группы в целом и обеспечивала определенный запас адаптаций к изменяющимся условиям среды. Тенденция к индивидуализации поведения прослеживается в сравнительном ряду приматов”141 .
Появление новой экологической ниши привело к тому, что при сортировке по поведенческим стратегиям “естественный отбор на “линии шимпанзе” стал усиливать качества, повышающие приспособленность к жизни в лесу небольшими коллективами, в то время как на линии гоминизации (очеловечивания) требовалось развитие таких качеств, как организация, сплоченность сравнительно большой группы, способной жить как в лесу, так и на границе саванны, более совершенные средства общения – коммуникации между членами группы, координация их действий”142 . “Комплекс гоминизации” (как целое) становится “объектом действия движущего естественного отбора, определяющего направление дальнейшего развития и видообразования”143 . У тех же, кто был лучше адаптирован для жизни в лесу, наоборот, развиваются специализации к лесной жизни – стопы, пригодные для лазанья по деревьям, руки, удобные не только для лазанья, но и для хождения с опорой на костяшки пальцев, зубная система, приспособленная к специфически лесной диете144 .
Первоначально между прежними и новыми биотопами шел постоянный обмен генетическим материалом (ср.: “сочетание интеграции на генетическом и дифференциации на экстрагенетическом уровнях позволяет популяциям одного вида приматов занимать различные экологические ниши и создает возможность для широкого расселения вида, освоения новых территорий, позволяет одновременно с этим сохранить видовое единство”145 ). Но климат становился все более холодным и сухим, саванны вытесняли леса, и постепенно территории, на которых жили гоминиды, отделялись друг от друга. Обмен особями (и, соответственно, генами) прекращался, и в разделившихся популяциях накапливались разные генетические изменения. У обитателей более лесных биотопов они были направлены на все лучшее приспособление к жизни в лесу (затем – в редколесье и т. д.) и соответствующему питанию, у обитателей более новых биотопов – на дальнейшее совершенствование поведения, развитие способности постигать причинно‑следственные связи и реагировать на события окружающей действительности “с опережением”. Соответственно, чем более гоминиды продвигались в саванны, тем больший спрос возникал на развитие коммуникативной системы.
Жизнь на границе более закрытого и более открытого ландшафта благоприятствует тому, чтобы особи, не забывая старых способов добычи пищи, поиска укрытий и т. п., осваивали новые. Количество потенциально возможных для вида поведенческих программ таким образом растет. Еще более оно возрастает с появлением орудий – помимо навыков, позволяющих добыть пищу при помощи орудий, полезно помнить навыки, позволяющие добыть ее “голыми руками”. Кроме того, полезно представлять себе, как эти орудия делаются, где добывается для них материал и т. д. Еще один фактор роста числа поведенческих моделей – переход к более активному употреблению мяса. Мясо стало важным источником энергии, и, следовательно, репертуар пищедобывательных стратегий расширился – независимо от того, какими конкретно способами это мясо добывалось (была ли это охота, поедание падали или же комбинация того и другого). При этом “не исключено, что изменение рациона и укрупнение мозга усиливали друг друга и способствовали развитию более сложного социального поведения, что вызывало дальнейшее совершенствование тактики пищевого поиска и диеты. А это, в свою очередь, благоприятствовало дальнейшей эволюции головного мозга”146 . С одной стороны, увеличивающийся мозг дает возможность поддерживать более развитую коммуникативную систему, с другой – появляется способность принимать во внимание все большее и большее количество факторов внешней среды, все тоньше различать детали, позволяющие отличить одну ситуацию от другой, и на развитие коммуникативной системы возникает спрос. Можно, конечно, этого и не делать, но тогда для выживания необходимо встроиться в более “традиционную” группировку – и продолжать существование до тех пор, пока старые местообитания еще остаются.
Процесс возникновения человеческого языка недоступен непосредственному наблюдению, поэтому судить о нем можно лишь по косвенным данным. В качестве одного из источников таких данных нередко называют развитие языка у ребенка. Стадии, выделяемые в овладении языком, представляются легко сопоставимыми либо с последовательно сменявшими друг друга видами гоминид, либо с палеолитическими индустриями. Основанием для этого служит так называемый “основной биогенетический закон” Эрнста Геккеля – онтогенез (т. е. развитие единичной особи) есть сжатое и сокращенное повторение хода филогенеза (т. е. развития таксона).
Но все же, по‑видимому, не следует прямо проецировать становление языка у ребенка на происхождение человеческого языка в целом. Биологам хорошо известно, что закон Геккеля представляет собой лишь частный, хотя и довольно широко распространенный случай. Он выполняется только для отдельных органов при их преобразовании по способу анаболии (т. е. добавления еще одной стадии развития после всех тех, что имелись у вида‑предка). Как было показано А.Н. Северцовым147 , эволюционируют не только фенотипы взрослых особей, но и их онтогенезы. Как пишет Н.Н. Иорданский, “изменчивость организмов проявляется на всех стадиях индивидуального развития и служит материалом для действия естественного отбора”148 . Онтогенез человека достаточно сильно изменился по сравнению с онтогенезом обезьян. Если детеныш обезьяны от рождения до взрослого состояния проходит две стадии – младенчество и детство, то у человека таких стадий осмысленно выделять четыре149 . Первая из них (от рождения до двух с половиной – трех лет) несколько перестроена по сравнению с обезьяньим младенчеством, вторая (до момента смены зубов) возникает в человеческой линии около 2 млн. лет назад150 (у обезьян этой стадии нет), третья (до начала пубертатного периода) соответствует детству (= подростковому возрасту) обезьян, но также претерпела значительные перестройки, и наконец, четвертая стадия (собственно подростковая, примерно до 19 лет), судя по палеонтологическим данным, появляется у неоантропов, а у Homo erectus и более ранних гоминид отсутствует151 . Эти периоды различаются не только по тому, какие телесные структуры формируются в течение каждого из них, но и по тому, какие изменения претерпевает когнитивная сфера, а также какими аспектами языка овладевает ребенок (см. гл. 1).
У человека, который усваивает язык, изначально (до появления речи) присутствует целый ряд необходимых для этого когнитивных установок, отсутствующих у обезьян, – стремление интерпретировать звуки, которые произносят другие люди, как знаки, желание узнавать названия различных объектов, установка на кооперативность в поведении вообще и в коммуникации в частности и т. д. (см. гл. 2). Соответственно, на каких‑то этапах эволюции человека они должны были войти в программу его онтогенеза. О перестройке программы индивидуального развития свидетельствует и то, что период овладения звуком (лепет) в ней предшествует периоду, когда ребенок начинает понимать, что окружающие имеют цели и намерения152 .
Еще одно весьма существенное отличие онтогенеза и филогенеза языка состоит в том, что человек выучивает язык в общении с теми, кто его уже знает (и это является необходимым условием, см. гл. 1; даже если родители – носители пиджина, все равно есть хотя бы один настоящий язык, которым они владеют в совершенстве, – их родной язык). Как показывают наблюдения психолингвистов, диалоги детей со взрослыми более чем вдвое (в среднем) длиннее, чем диалоги детей с детьми (лучше всего детям удается поддерживать беседу, когда они вступают в разговор взрослого с ребенком на пару лет старше их, – в этом случае их реплики не сбивают разговор с обсуждаемой темы, а самих реплик может быть больше, чем обычно бывает в разговоре с этим же взрослым или с этим же ребенком)153 .
Дата добавления: 2016-02-02; просмотров: 738;