Как получить новое, унаследовав старое? 1 страница
Человекообразные обезьяны в языковых проектах демонстрируют удивительные способности, но все же овладеть языком на уровне взрослого человека оказываются не в состоянии. Не демонстрируют они сравнимых с человеческими языковых способностей и в природе. Это значит, что наши общие с обезьянами предки еще не имели языка. Откуда же взялся язык, если его раньше не было? Человек, как известно, “произошел от обезьяны” {33}. Значит, человеческий язык каким‑то образом получился в результате эволюции из коммуникативной системы наших “обезьяньих” предков.
Еще до недавних пор многие специалисты представляли себе это эволюционное развитие примерно так: у кого‑то из предков человека появилась генетическая мутация, в результате которой он обрел языковую способность1 . На это как будто бы указывают данные специфического нарушения речи (англ. SLI – specific language impairment ), обусловленные повреждением гена FOXP 2 на седьмой хромосоме2 . Люди, страдающие этим наследственным заболеванием, испытывают трудности с грамматикой своего родного языка – примерно такие же, которые многие из обычных людей испытывают с грамматикой иностранного. Они не чувствуют некоторых различий между регулярной и нерегулярной морфологией, иногда оказываются не в состоянии построить по общему правилу форму от незнакомого слова. С. Пинкер3 , опираясь на исследования Мирны Гопник, приводит случай, когда женщина, страдающая SLI , не справилась с так называемым ваг ‑тестом {34}: человеку показывают изображение некоторого вымышленного существа и говорят, что оно называется ваг (англ. wug ), а потом показывают картинку с двумя такими существами и просят закончить фразу Здесь два … Для обычного четырехлетнего носителя английского языка не составляет никакого труда сказать, что на картинке изображены two wugs , но человеку, страдающему SLI , нелегко сообразить, какое правило надо применить в данном случае. Как показывают исследования Татьяны Владимировны Черниговской и Киры Гор4 , люди, страдающие SLI , как бы хранят формы слов в памяти в целом виде, почти не проводя их морфологического анализа. Возможно, похожим образом обстоит дело и с предложениями: понимание конструкций с необычным порядком слов вызывает у таких больных значительные трудности, “грамматические процедуры как бы симулируются за счет памяти и эксплицитных правил”5 . Нарушается при SLI и фонетический уровень: как пишет Черниговская, у таких людей отмечаются “монотонная речь, нарушение речевого ритма и неправильное членение звукового потока; нарушение просодики, ударения как в частотных, так и в незнакомых, но ясных с точки зрения правил данного языка, словах; сложности произнесения многосложных слов (сокращение их за счет пропуска слогов)”6 . Кроме того, у них несколько нарушен моторный контроль за мышцами рта – им сложно, например, высунуть язык или дунуть по команде7 .
Как показывают исследования, ген FOXP 2 подвергся мутации в ходе эволюции человека: две из трех замен, происшедших в этом гене со времени существования общего предка человека и мыши, произошли после разделения гоминид и человекообразных обезьян9 .
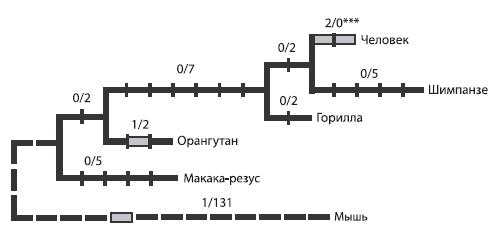
Рис. 5.1. Эволюция гена FOXP 28 . Цифра до знака дроби обозначает количество значимых нуклеотидных замен, после знака дроби – число незначимых (“синонимичных”), то есть не приводящих к синтезу другой аминокислоты замен.
Не означает ли это, что причиной возникновения человеческого языка послужила одна‑единственная макромутация, сформировавшая речевой аппарат, изменившая форму черепа, перестроившая мозг и “вложившая” в него языковую способность, как утверждали авторы некоторых работ10 ? Так, по мнению Д. Бикертона, такая мутация вполне могла бы произойти у “митохондриальной Евы”. А Тимоти Кроу, напротив, связывает языковую мутацию с Y‑хромосомой, наследуемой исключительно по мужской линии {35}.
Все же, по‑видимому, идея о том, что человеческий язык мог возникнуть в результате единственной мутации, неверна (показательно, что в своей более поздней работе Д. Бикертон от нее отказывается11 ). В гл. 2 мы видели, сколько различных механизмов – как анатомо‑физиологических, так и когнитивных – должны слаженно работать, чтобы люди могли пользоваться языком. Органы, задействованные в обеспечении функционирования языка, возникают в процессе развития из разных эмбриональных зачатков – например, в развитии слухового анализатора (весьма ценного для звучащей речи) участвуют слуховая плакода, зачаток рудиментарной жаберной щели, элементы подъязычной и челюстной висцеральных дуг, мезодерма головы и невральная эктодерма12 . И это лишь один из множества необходимых для работы языка компонентов. Как пишет биолог‑эволюционист Николай Николаевич Иорданский, “совершенно ясно, что невозможно возникновение всей этой сложной, но целостной системы как путем случайного подбора различных мутаций (например, при дрейфе генов), так и посредством одной крупной “системной” мутации”13 . Очевидно, что перестройка речевого аппарата и формирование нужных мозговых структур и всех прочих, столь многочисленных, сложно устроенных и хорошо сбалансированных приспособлений в результате случайной замены нуклеотидов в молекуле ДНК не более вероятны, чем сборка самолета смерчем, пронесшимся по свалке14 .
На это иногда возражают, что генетические изменения имеют так называемый “плейотропный” эффект – мутация, происшедшая в одном гене, может приводить к множественным последствиям в самых разных структурах организма. Но приводимые в литературе примеры плейотропного эффекта – это примеры разрушений. И действительно, нарушить работу сложного механизма, заменив в нем даже всего одну деталь на негодную, тем легче, чем сложнее механизм, но получить слаженно работающую систему, добавив “лишнюю” деталь, гораздо труднее – это понимает всякий, кто пытался если не переделать собственными руками велосипед в мотоцикл, то хотя бы установить на свой компьютер программу, написанную для другой операционной системы.

Рис. 5.2. Разрушить такое сооружение, вытащив любую карту из нижнего яруса, очень легко, а вот создать нечто подобное, подложив карту в нижнюю часть кучи карт, невозможно.
Муха‑дрозофила, получившая в результате мутации лапку на месте усика‑антенны или четыре крыла вместо обычных двух (см. фото 18 на вклейке), безусловно, является другим существом, сильно отличающимся от своих родителей (в последнем случае это различие на уровне отрядов). Но никакой выгоды для мухи в этом нет: весь ее организм рассчитан на два крыла, пару жужжалец {36}, две антенны и шесть лапок на своих местах. Такого рода мутации не создают ни систем управления, которые бы позволяли “лишним деталям” слаженно взаимодействовать с прочими частями организма, ни поведенческих программ, которые бы обеспечивали эффективное использование новообретенных признаков15 .
Против идеи единственной макромутации говорит и то, что генов, так или иначе принимающих участие в обеспечении функционирования языка и подвергшихся изменениям в ходе эволюции человека, обнаруживается с каждым годом все больше16 . Таков, например, ген, кодирующий альфа‑текторин (один из важных компонентов текториальной мембраны внутреннего уха)17 . В той же статье в журнале “Science” , написанной большой группой авторов, перечисляются и другие гены, претерпевшие изменения в ходе развития человеческого слухового анализатора, – DIAPH1, FOXI1, EYA4, EYA1 и OTOR . Явно служил мишенью отбора в гоминидной линии ген HAR1 (точнее, это не ген, а участок гена HAR1F , кодирующего регуляторную РНК), – человек и шимпанзе имеют в нем 118 отличий, тогда как шимпанзе и курица – всего два18 . Ген FOXB1 необходим для обеспечения оперативной памяти – а при сбоях в оперативной памяти человек не сможет полноценно пользоваться языком (например, он не сможет обобщить синтаксические правила, определенные на достаточно длинных предложениях). Определенную роль в овладении языком играет ген c‑fos 19 , который включается в мозге в ответ на новизну (т. е. в том случае, когда поступающие в организм внешние стимулы не находят соответствия в том, что уже хранится в памяти). Важны для языка и гены MCPH1 20 и ASPM 21 , регулирующие объем мозга, и многие другие гены – все те, которые принимают участие в обеспечении механизмов памяти, планирования поведенческих программ, генерализации правил, формировании в ходе развития организма (в том числе и после рождения) необходимых анатомических структур и т. д. Можно, по‑видимому, утверждать, что язык, как и многие поведенческие признаки, имеет полигенное наследование22 .
Кроме того, гены в принципе не могут кодировать грамматические правила – каждый ген является единицей транскрипции, отрезком ДНК, на базе которого синтезируется одна молекула РНК (информационная, рибосомная, транспортная или регуляторная). Записать на него программу типа “ставь подлежащее перед сказуемым” или “ставь подлежащее после сказуемого” (с тем чтобы потом можно было “перещелкнуть переключатель” между этими двумя возможными вариантами), по‑видимому, технически невозможно.
Соответственно, присущая человеку “врожденная языковая способность” предстает не как набор предзаданных правил Универсальной Грамматики, а как стремление искать структуру и “заполнять пустые клетки” в логически – по презумпции – организованной “таблице” коммуникативной системы. Авторы многих работ признают, что врожденными являются не свойства грамматики языка, а свойства механизма его усвоения23 , не детали коммуникативной системы, а “механизмы их выведения из поведения окружающих”24 . В книге Р. Джакендоффа25 предпринимается попытка совместить идею врожденности УГ с уже не подлежащим сомнению фактом отсутствия специального “языкового органа” (который должен был бы, согласно гипотезе Н. Хомского, ее содержать). Универсальная Грамматика в интерпретации Р. Джакендоффа не является “монолитной способностью”, а состоит из нескольких автономных слоев26 , представляя собой не набор правил, а “устройство для овладения языком” (англ. LAD – Language Acquisition Device ). При этом наследуется УГ тем же способом, что и поведение.
Наличие универсальных закономерностей (как в области грамматики, так и в области “универсального словаря” – “мыслекода”) объясняется, вероятно, тем фактом, что у всех людей много общих перцептивных, поведенческих, эмоциональных предрасположенностей27 , а даже слабые предрасположенности, если они есть всегда, ведут к значительным последствиям28 . Следует, впрочем, отметить, что универсальность этих закономерностей относительна: хотя они представлены во многих языках, но едва ли не для каждой из них можно найти такой язык, в котором она отсутствует29 .
Что же касается мутации в гене FOXP2 , то она в любом случае произошла задолго до “митохондриальной Евы” – такой же, как у нас, вариант этого гена был обнаружен у неандертальца30 .
Вообще, следует отметить, что от гена и мутации в нем до внешнего (или, как говорят биологи, фенотипического) свойства – длинный путь. В клетке существуют специальные ферменты, которые исправляют ошибки, возникающие при копировании ДНК (так что в конечном счете остается примерно 1 ошибка на 10 млн. пар нуклеотидов)31 . Многие из остающихся замен синонимичны: например, если в последовательности ЦЦТ заменить последний нуклеотид Т на А, Ц или Г, полученный триплет все равно будет кодировать ту же самую аминокислоту – глицин. Замена одной аминокислоты в белке может никак не повлиять на его функцию. Но даже в том случае, если мутация окажется значимой, другие гены, имеющиеся у организма, могут усилить или ослабить ее действие и даже полностью свести его на нет. Так, например, гиляцкие лайки – совершенно бесстрашные собаки, несмотря на то, что генетически они предрасположены к трусости. Их мало кому доводилось видеть испуганными, поскольку эти лайки очень маловозбудимы32 – их, как говорится, не проймешь. Генетические характеристики, обусловливающие малую возбудимость, блокируют проявление трусости. Таким образом, даже если предположить, что одна мутация вызвала у человека некую “предрасположенность” к языку, в отсутствие множества других изменений эта предрасположенность с большой вероятностью вовсе никак бы не проявилась.
У видов, использующих исключительно половое размножение, в каждом следующем поколении набор генов, которые могут влиять на внешнее проявление свойства, связанного с мутантным геном, будет изменяться, поскольку половина генетического материала достается организму от одного из родителей, а половина – от другого. И проявиться в виде фенотипического свойства может лишь та мутация, которой не помешает в этом вторая половина генома. Поэтому, например, у женщин – носительниц гемофилии внешних признаков этого заболевания не наблюдается: работа второй X‑хромосомы, не затронутой мутацией, обеспечивает им в конечном итоге вполне удовлетворительный уровень свертываемости крови.
Усложняет связь между генами и фенотипическими характеристиками и тот факт, что новые свойства, отсутствовавшие у родителей, организм может получить не только в результате мутаций, но и в результате ненаследственных изменений – модификаций или морфозов. Например, попадание некоторых веществ в организм вполне генетически нормальной беременной женщины может повлечь развитие у плода врожденных уродств33 . Дрозофила с четырьмя крыльями вместо двух может появиться не только вследствие мутации (вернее, двух мутаций – bithorax и postbithorax ), но и том в случае, если на ранней личиночной стадии она попадет под воздействие эфира или высокой температуры34 . Такое изменение не будет наследоваться, оно будет, как говорят биологи, фенокопией данной мутации (а сама мутация, соответственно, генокопией данного морфоза). Между внешними проявлениями мутаций и модификаций существует, как показал биолог‑эволюционист Иван Иванович Шмальгаузен, глубокий параллелизм35 , основанный на том, что набор белков, которые может синтезировать клетка, далеко не безграничен: чаще всего речь идет о том, что характерные для организма белки начинают синтезироваться с другой интенсивностью или в другие сроки, и именно этим определяется набор потенциально возможных отклонений от исходного фенотипа.
Роль внешней среды для формирования тех или иных внешних признаков нередко оказывается весьма существенной даже для тех организмов, которые обладают значительной автономностью развития: существуют некоторые элементы среды, которые полезно “заметить” для повышения приспособленности, и естественный отбор благоприятствует тем организмам, в чьем развитии заложена программа реагировать на них определенным образом. Например, такое свойство, как способность к импринтингу {37}, полезно, поскольку оно позволяет птенцу выработать эффективную реакцию на конкретную особь – собственного родителя. Более того, в индивидуальном развитии (онтогенезе) существуют специальные чувствительные периоды, когда развивающийся организм наиболее восприимчив к воздействиям определенного типа. Есть чувствительный период и в усвоении языка (см. гл. 1). Если человек не овладел языком в этот период, он, скорее всего, уже не сможет стать полноценным носителем; “вероятные причины этого – возрастные изменения в мозге, такие как уменьшение уровня метаболизма и количества нейронов на протяжении младшего школьного возраста и достижение наименьшего количества синапсов и уровня метаболизма примерно в пубертатном возрасте”36 .
Внешние по отношению к организму факторы могут существенно изменить проявления того, что закодировано в его генах. Например, добиться “пассивно‑оборонительной” (по терминологии специалиста по поведению животных Леонида Викторовича Крушинского), т. е. трусливой, реакции от гиляцких лаек можно, если искусственно поднять уровень их возбудимости путем введения кокаина37 . Под действием факторов внешней среды может изменяться в том числе устройство мозга – как уже говорилось в гл. 2, нейроны его коры могут перепрофилироваться. Если у младенца, еще не овладевшего языком, удалить все левое полушарие (такое иногда приходится делать по медицинским показаниям), он сможет впоследствии научиться говорить – языковые центры разовьются в сохранившемся правом38 .
Более того, различные мутации при определенном подборе разных условий могут вызывать одинаковые внешние проявления. При изучении поведения лабораторных мышей было выяснено, что, например, разницу в уровне исследовательской активности у мышей двух различных линий можно нивелировать (и даже заменить на противоположную) путем введения некоторых фармакологических веществ39 . Роберт Трайон вывел линии “умных” и “глупых” крыс – первые значительно быстрее, а вторые значительно медленнее обучаются находить пищу в 17‑тупиковом лабиринте. Но в 16‑тупиковом лабиринте крысы обеих линий демонстрируют одинаковые успехи40 .
Таким образом, то, каким в итоге окажется человек (в том числе в аспекте владения языком), зависит от генов его родителей и их конкретной комбинации (например, от родителя, имеющего генетическую предрасположенность к SLI , ребенку может передаться не дефектный, а второй, нормальный вариант гена FOXP2 ), от характеристик цитоплазмы яйцеклетки (так называемый “материнский эффект” в узком смысле), от условий внутриутробного развития (например, употребление матерью алкоголя может крайне негативно сказаться на развитии мозга плода, что может помешать ему впоследствии успешно овладеть языком) и, наконец, от тех воздействий, которые окажет на него окружающая среда после рождения (например, сильный испуг может сделать человека заикой). Именно поэтому, кстати, при клонировании невозможно получить точную копию родителя – для этого пришлось бы воспроизвести в точности не только гены, но и все релевантные условия, регулирующие индивидуальное развитие. И именно поэтому, сколь бы умны ни были родители человека, сколь бы хорошо они ни владели языком, это не дает стопроцентной гарантии, что их потомки будут умными и красноречивыми.
Разумеется, гены в наследовании играют очень важную роль. Во‑первых, они задают возможный спектр изменчивости: какие свойства можно получить в результате мутаций, а какие – нельзя. Например, невозможно получить синие глаза у дрозофилы (по крайней мере, среди изученных примерно двух миллиардов дрозофил синеглазые особи не попались ни разу). Не существует мутаций, которые бы случайным образом наделяли любые организмы любыми свойствами41 так, чтобы из них потом можно было отбирать оптимальные, – и в этом ответ на знаменитый вопрос “Почему у свиней нет крыльев?”42 . Их нет не потому, что естественный отбор отбраковал всех крылатых свиней, появившихся в результате мутации, а просто потому, что набор генов, характерных для свиней (как и для всех других копытных), не предусматривает возможности появления крыльев – ни в качестве дополнительной пары конечностей, ни в качестве модификации уже имеющихся.
Во‑вторых, гены определяют, на какие факторы внешней среды организм (в том числе в своем развитии) будет реагировать в большей, а на какие – в меньшей (в частности, в нулевой) степени и каков будет характер этой реакции. В‑третьих, под генетическим контролем находится то, в какой период развития тот или иной фактор внешней среды будет оказывать сильное (подчас определяющее) воздействие на организм, а в какой – слабое (или вообще никакого). Общий генофонд популяции задает частоту, с которой будут возникать те или иные характеристики особей (как анатомо‑физиологические, так и поведенческие), и даже частоту появления различных мутаций. В число факторов среды, оказывающих влияние на развитие организма на любой отдельно взятой стадии, входят результаты работы генов на стадии предшествующей.

Рис. 5.3. Чтобы дети выросли заботливыми, их надо как следует вылизывать.
Еще один существенный для понимания механизмов наследования момент состоит в том, что работа генов не заканчивается с рождением организма, и внешние условия могут оказывать на нее влияние. Например, как было выяснено недавно, не только гены могут управлять поведением, но и, наоборот, поведение может управлять генами43 . Американские исследователи Джин Робинсон, Рассел Ферналд и Дэвид Клейтон задались вопросом о механизме наследования родительской заботы у крыс. Оказалось, что этот феномен определяется не родительскими генами и не научением. Просто, когда крыса‑мать облизывает своих детенышей, у них происходит метилирование определенных участков ДНК, что вызывает “выключение” ряда генов, и в результате изменяется реакция нейронов на некоторые гормоны, а это, в свою очередь, приводит в дальнейшем к увеличению уровня заботы выросших крысят о своих собственных детенышах. Метилирование ДНК – не генетический (а так называемый “эпигенетический”) процесс, но при нормальном ходе событий (крыса‑мать вылизывает своих детенышей, из них вырастают заботливые матери, которые, в свою очередь, вылизывают своих детенышей и т. д.) это обеспечивает вполне эффективное наследование данного поведенческого признака44 .
Все это показывает, что попытки однозначно определить, что же все‑таки сыграло ключевую роль в возникновении языка – гены или среда (проблема, получившая в англоязычной литературе название “nature– nurture ”, “природа – воспитание”), едва ли могут привести к осмысленному результату. Скорее, на мой взгляд, разумно изучать взаимодействие генов и среды в этом процессе.
Эволюция живых организмов происходит путем естественного отбора. Но естественный отбор не работает с мутациями напрямую – ему доступны лишь фенотипы. Так, если бы дрозофилам по какой‑то причине стало выгодно иметь четыре крыла, в выигрыше оказались бы не только носители соответствующего комплекса мутаций, но и те, кто получил дополнительную пару крыльев в результате морфоза (хотя, разумеется, последние не смогли бы передать эту особенность своим потомкам).
Новые фенотипические характеристики могут возникнуть не только непосредственно в результате мутаций, но и вследствие комбинационной изменчивости – гены родителей, работая вместе в одном организме, могут наделять его тем или иным свойством, которого не было в фенотипе ни у одного из родителей. Например, как показал Л.В. Крушинский45 , если скрестить бесстрашную немецкую овчарку с не менее бесстрашной гиляцкой лайкой, получится очень трусливая собака, поскольку щенок унаследует от родителя‑лайки склонность к пассивно‑оборонительной реакции, а от родителя‑овчарки – высокую возбудимость. В каждой популяции циркулирует определенное количество вариантов (аллелей) различных генов, и признаки, получаемые в результате их комбинаций, хотя и не наследуются напрямую, тем не менее возникают у членов этой популяции вновь и вновь. Частота появления особей с теми или иными фенотипическими особенностями входит в набор характеристик вида (и может меняться со временем).
Для успешного прохождения отбора не имеет значения не только то, каковы в точности у особи гены, но и то, каковы ее отдельные свойства, – необходимо лишь, чтобы весь комплекс присущих особи индивидуальных признаков в комбинации с характеристиками той среды, где она живет, обеспечивал наилучшие показатели выживания и размножения. Например, в районах, где распространена малярия, у многих людей в генах заложена устойчивость к ней. Как выяснили генетики, обеспечивается она несколькими различными мутациями (а значит, формировалась в разных регионах независимо)46 . При этом сама по себе особенность, кодируемая соответствующими генами, вредна – человек, получивший мутантные гены от обоих родителей, быстро умирает от серповидно‑клеточной анемии. Однако в условиях значительной угрозы малярии плюс возможности комбинации с немутантными генами второго родителя такая мутация поддерживается естественным отбором. (Заметим в скобках, что отбор в данном случае закрепляет не только саму мутацию, но и уровень ее распространения – ненулевой, но и не стопроцентный.)
Любое отдельное свойство может быть в разных условиях и положительным, и отрицательным, и нейтральным. Например, густой волосяной покров для млекопитающих, живущих в холодном климате, является благом, поскольку защищает их от потери тепла, но для земноводных с их кожным дыханием или, скажем, для бегемотов, которым крайне важно избегать перегрева, был бы смертелен. Склонность издавать громкие звуки при виде опасности повышает выживаемость у общественных животных, но понизила бы ее у животных, ведущих одиночный образ жизни.
Поэтому при возникновении новых фенотипических особенностей – как вследствие мутаций, так и в результате модификаций и комбинирования родительских генов – лучше приспособленным оказывается не тот организм, у которого соответствующие показатели выражены в наиболее сильной степени, а тот, который смог наиболее успешно “вписать” их в контекст как своих собственных характеристик, так и условий окружающей среды.
У каждого вида есть так называемая “норма реакции” – это, по определению теоретика эволюции Алексея Сергеевича Северцова, “пределы, в которых может изменяться фенотип без изменения генотипа”47 ; в этих пределах организмы могут приспосабливаться к условиям среды. Когда какие‑то фенотипические черты (возникающие в результате комбинации родительских генов, или реакции на какие‑то факторы внешней среды, или и того, и другого), а точнее комплексы черт, в определенных условиях оказываются полезными, организмы, их имеющие, лучше выживают и размножаются, и отбор благоприятствует тем, в чьем потомстве эти свойства проявляются с большей вероятностью (и большей надежностью). Соответственно, рано или поздно вырабатывается механизм, с достаточной надежностью обеспечивающий совместное наследование удачно сочетающихся между собой признаков – возникают мутации, повышающие вероятность передачи следующему поколению всего этого комплекса целиком48 (в пределе – до единицы), и эти мутации закрепляются отбором.
Подобные мутации могут происходить в генах, которые регулируют активность других генов. И, вероятно, не случайно, что многие генетические отличия человека от шимпанзе связаны именно с имеющейся в геноме системой регуляции. Так, например, американский генетик Грегори Рэй и его коллеги недавно обнаружили49 , что у нас имеется мутация в регуляторной области гена, кодирующего белок продинорфин. Из этого белка в организме образуются эндорфины – вещества, участвующие в регуляции эмоций, влияющих на социальное поведение, формирование привязанностей, способности к обучению. Сам белок продинорфин у человека и шимпанзе идентичен, но у человека его синтезируется несколько больше (примерно на 20 %), а кроме того, по‑видимому, различаются реакции на внешние стимулы – в каких случаях продинорфина будет синтезироваться больше (и насколько), а в каких – меньше. А это, как пишет А.В. Марков50 , “могло серьезно изменить мотивацию человеческих поступков, наши желания и “жизненные цели””. Группа молекулярных биологов из США и Австралии, сравнив уровень активности 907 генов у человека и нескольких видов обезьян, обнаружила, что многие гены в ходе эволюции разных групп приматов увеличили или уменьшили свою активность. При этом среди генов, активность которых в человеческой линии повысилась, оказалось много таких, которые участвуют в регуляции активности других генов51 . Регулятором (причем регулятором высокого уровня – т. е. таким, который регулирует активность других генов‑регуляторов) оказался и знаменитый ген FOXP25 2 . Начинающийся с него “генетический каскад отвечает за нормальное развитие мозга, в особенности тех отделов, от которых зависит координация движений и, в том числе, артикуляция речи”53 .
В любой популяции за счет комбинирования генов (обеспечиваемого половым размножением) и воздействия окружающей среды появляются различные фенотипы с различной частотностью. Если какой‑то фенотип (именно фенотип в целом, обладающий некоторым балансом свойств) оказывается очень удачным, то преимущество получит та популяция, где такой фенотип (вне зависимости от его генетической основы) будет появляться чаще. И если перемещение и скрещивание особей внутри этой популяции происходит существенно чаще, чем между этой популяцией и соседними, то популяция, где чаще появляется удачный фенотип, будет производить большее количество потомства и, соответственно, “экспортировать” особей в другие популяции. Даже если среди этих особей будет не очень много носителей удачного фенотипа, принесенный ими генетический “багаж” будет способствовать тому, чтобы удачный фенотип и на новом месте появлялся с большей частотностью. И таким образом генофонд, способствующий более частому появлению удачного фенотипа, будет распространяться.
Дата добавления: 2016-02-02; просмотров: 798;