Коммуникация в мире животных
Исследование происхождения человеческого языка невозможно без изучения коммуникативных систем животных – иначе мы не сможем выделить ни то новое, что появилось у человека по сравнению с животными, ни те полезные для развития языка свойства, которые к началу его эволюции уже имелись. Неучет факторов такого рода ослабляет выдвигаемые гипотезы. Например, Т. Дикон отводит ключевую роль в происхождении языка употреблению знаков‑символов (его книга так и называется – “The symbolic species”, “Символический вид”1 ) – но поскольку способность к их употреблению обнаруживают и многие животные (причем, как мы увидим ниже, не только в условиях эксперимента), на роль главной движущей силы глоттогенеза пользование символами не годится.
Впрочем, исследование коммуникации животных нужно не только для того, чтобы отвергать подобные гипотезы. Нынешнее состояние науки позволяет поставить и более глубокие вопросы: с чем коррелирует наличие у коммуникативной системы тех или иных характеристик? Какие существуют направления эволюции коммуникативных систем и чем они могут определяться?
Прежде всего необходимо понимать, что за словом “животные” скрывается огромное количество самых разных существ, одни из которых близки к человеку до такой степени, что осмысленно ставить вопрос о тех свойствах, необходимых для коммуникации, которыми обладал их общий предок, другие же далеки настолько, что у общих предков заведомо никаких релевантных для коммуникации свойств быть не могло. Таким образом, следует различать “гомологии” и “аналогии” – под первым термином понимаются свойства, развившиеся из того общего наследия, которое досталось от общего предка, под вторым – характеристики, которые, будучи внешне сходными, развились в ходе эволюции независимо. Например, наличие двух пар конечностей у человека и крокодила – гомология, а обтекаемая форма тела у рыб, дельфинов и ихтиозавров имеет аналогическую природу.
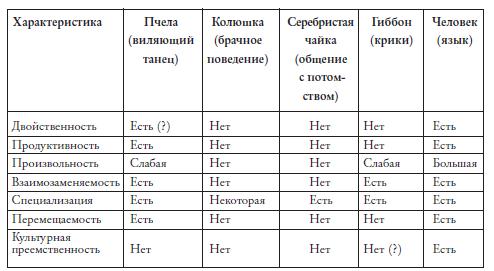
Рис. 4.1. Сравнение языка с коммуникативными системами других видов по критериям Ч. Хоккета 2 .
Когда по критериям, предложенным Ч. Хоккетом, было проведено сравнение языка с коммуникативными системами нескольких разных видов животных (колюшки, серебристой чайки, пчелы и гиббона), оказалось, что больше всего общих черт с языком набирает коммуникативная система медоносной пчелы (Apis mellifera ). Виляющий танец пчел обладает такими свойствами, как продуктивность и перемещаемость; он является специализированным коммуникативным действием; те, кто может производить сигналы этого типа, могут и понимать их (последнее называется “свойством взаимозаменяемости”). До некоторой степени в танце пчел можно усмотреть даже произвольность знака: один и тот же элемент виляющего танца у немецкой пчелы обозначает расстояние в 75 метров до источника корма, у итальянской – 25 метров, а у пчелы из Египта – всего пять3 . Соответственно, эта коммуникативная система является (по крайней мере, отчасти) выучиваемой – как показали эксперименты Нины Георгиевны Лопатиной4 , пчела, выращенная в изоляции и не имевшая возможности наблюдать за танцами взрослых особей, не понимает смысла танца, не может “считывать” с него передаваемую информацию. С формальной точки зрения в танцах пчел можно выделить элементарные компоненты (см. ниже), различные комбинации которых составляют разные смыслы (подобно тому, как в человеческом языке различные комбинации фонем дают разные слова)5 .
Определенные аналогии можно усмотреть между человеческим языком и коммуникативными системами некоторых видов муравьев. Как показали опыты Ж.И. Резниковой (см. фото 16 на вклейке), проведенные с муравьями‑древоточцами Camponotus herculeanus , их сигнализация обладает свойством продуктивности и свойством перемещаемости: муравьи способны сообщать своим сородичам о различных местах нахождения корма. При этом они могут сжимать информацию: путь типа “все время направо” описывается короче, чем путь типа “налево, потом направо, еще раз направо, потом налево и потом направо”. Информация о том же самом, хорошо знакомом месте передается быстрее, чем о другом. Хотя прямой расшифровке коммуникативная система муравьев не поддается, эта аналогия показывает, что такие свойства, видимо, с неизбежностью возникают в коммуникативной системе, которая должна обеспечивать передачу большого количества разнообразной информации.
Как отмечает Ж.И. Резникова, использование разными видами муравьев разных типов передачи информации связано с их образом жизни и теми задачами, которые им приходится решать. Тем видам, у которых численность семьи составляет не более нескольких сотен особей, развитая знаковая система не нужна: необходимое количество корма вполне можно собрать на расстоянии двух‑трех метров от гнезда, “а на таком расстоянии прекрасно действует и пахучий след”6 . Напротив, у тех видов, которые живут огромными семьями и собирают корм, удаляясь от гнезда на значительное расстояние, имеются коммуникативные системы, обладающие богатыми выразительными возможностями.
Для звучащей речи большое значение имеют формантные различия – прежде всего именно по ним (а не, скажем, по громкости, длительности или высоте основного тона) мы отличаем разные фонемы друг от друга. Но способность использовать формантные различия представлена и у животных. Как свидетельствует Т. Фитч, виды, использующие звуковую коммуникацию, – например, зеленые мартышки (верветки), японские макаки, журавли, – способны различать форманты не хуже людей7 . Даже у лягушек есть специальные детекторы, настроенные на те частоты, которые особенно важны для каждого конкретного вида. Формантные различия могут использоваться, в частности, для того, чтобы отличать друг от друга сородичей8 , для распознавания разных типов сигналов опасности и т. п.
Множество аналогов в животном мире имеет человеческая способность к рекурсии. Самый простой (по крайней мере, с точки зрения человека) мыслительный процесс, требующий применения рекурсии, – это счет: каждое следующее число на единицу больше предыдущего. Но считать, как показали исследования, умеют не только люди9 , но и шимпанзе (этому посвящены, в частности, специальные эксперименты, проводимые в Киото под руководством Тецуро Мацузавы10 ), попугаи11 , ворóны12 и муравьи13 . В опытах З.А. Зориной и А.А. Смирновой было показано, что серые вороны могут складывать числа в пределах 4 (и даже оперировать при этом обычными “арабскими” цифрами), муравьи в экспериментах Ж.И. Резниковой продемонстрировали способность “складывать и вычитать в пределах 5”14 . Макаки‑резусы (в опытах американских исследователей Элизабет Бреннон и Герберта Террейса) “считали” (последовательно дотрагиваясь на экране до изображений групп с разным количеством предметов) по возрастанию и по убыванию от 1 до 4 и от 5 до 915 .
Наиболее разработана аналогия между человеческим языком и песней певчих птиц (это один из подотрядов отряда воробьиных). Песня делится на слоги – отдельные спектральные события, имеющие более звучную вершину и менее звучные края. Каждый отдельный слог, подобно фонеме, не имеет собственного значения {28}, но их последовательность складывается в песню, несущую определенный смысл. Для распознавания песни существенно, чтобы слоги шли в определенном порядке – иначе представители соответствующего вида не опознают песню как свою16 .
Подобно языку, песня выучивается во время чувствительного периода, т. е. в ее передаче велико значение культурной составляющей. В чувствительном периоде есть стадия “лепета” (или “подпесни”, англ. subsong ) – подросший птенец‑слеток издает разнообразные звуки, как бы пробуя различные возможности голосового аппарата17 . Издает, в отличие от взрослых самцов, негромко, что называется, “себе под нос”. Для нормального развития вокального репертуара ему необходимо слышать и самого себя, и взрослых представителей своего вида. Обучение происходит посредством звукоподражания, причем это подражание является самоподдерживающимся – как и детям, овладевающим языком, птенцам не нужно специальное поощрение за выученные элементы коммуникативной системы. В результате такого обучения складываются – как и в языке – диалекты (местные варианты песни) и идиолекты (индивидуальные варианты песни, которые в работах орнитологов также именуются “диалектами”, что создает некоторую путаницу). У птиц имеется латерализация мозга, причем звукопроизводством управляет в нормальном случае левое полушарие.
Рис. 4.2. Сонограмма песни зяблика (Fringilla coelebs).
У певчих птиц, а также у попугаев и колибри, которые тоже обучаются своим звуковым коммуникативным сигналам посредством звукового подражания, контроль за звукопроизводством осуществляется не теми мозговыми структурами, что у тех видов, у которых звуковые сигналы являются врожденными18 . Повреждения аналогичных участков мозга приводят к аналогичным нарушениям звукопроизводства: при одних птицы, подобно людям с афазией Брока, теряют способность правильно составлять последовательности звуков, при других – способность выучивать новые звучания, при третьих – сохраняют лишь способность к эхолалическому повторению19 .
Множество аналогичных черт у языка и с коммуникацией китообразных. В обоих случаях носителем информации является звук (правда, у китообразных, в отличие от человека, большая часть сигналов передается в ультразвуковом диапазоне). У дельфинов есть “имена собственные” – знаменитый “свист‑подпись”: этим сигналом (индивидуальным для каждой особи) дельфины завершают свои сообщения, и с его помощью их можно позвать. У касаток Orcinus orca были обнаружены локальные диалекты20 . Как и в языках людей, одни “слова” (звуковые сигналы) у касаток более стабильны, другие сравнительно быстро (у касаток – на протяжении порядка 10 лет) меняются21 .
Звуковые сигналы дельфинов‑афалин (Tursiops truncatus ), согласно наблюдениям В.И. Маркова22 , комбинируются в комплексы нескольких уровней сложности. Комплекс, состоящий из нескольких звуков, сгруппированных определенным образом, может входить составной частью в комплекс более высокого уровня подобно тому, как слово, состоящее из нескольких фонем, входит составной частью в более сложный комплекс – предложение. Так же, как фонема может быть описана как совокупность смыслоразличительных признаков, в звуковых сигналах дельфинов могут быть выделены отдельные компоненты, противопоставляющие один звук другому.
Скорее всего, столь сложное устройство сигналов говорит о том, что у дельфинов (как и у людей) есть возможность (а значит, вероятно, и необходимость) кодировать большое (по подсчетам Маркова, потенциально даже бесконечно большое) количество разнообразной информации.
По‑видимому, коммуникативная система дельфинов позволяет им передавать в том числе и весьма конкретные сведения. В эксперименте, проведенном Уильямом Эвансом и Джарвисом Бастианом23 , два дельфина (самец Базз и самка Дорис) были обучены нажимать на педали в определенном порядке, чтобы получать пищевое подкрепление. Порядок менялся в зависимости от того, ровно горела лампочка над бассейном или же мигала, а подкрепление выдавалось лишь в том случае, когда на педали в правильном порядке нажимали оба дельфина. Когда лампочку установили так, чтобы ее могла видеть только Дорис, она оказалась в состоянии “объяснить” Баззу через непрозрачную стенку бассейна, в каком порядке следует нажимать на педали, – в 90 % случаев правильно.
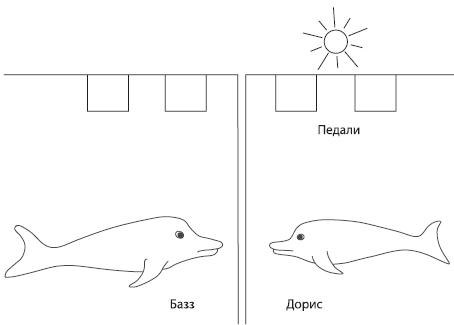
Рис. 4.3. Схема опыта В. Эванса и Дж. Бастиана 2
В опытах В.И. Маркова и его коллег дельфины передавали друг другу информацию о размере мяча (большой он или маленький) и о том, с какой стороны предъявляет его экспериментатор (справа или слева)25 .
Как показали Дэвид и Мелба Колдуэллы, дельфины, подобно людям, способны опознавать сородичей по голосу – независимо от того, что конкретно тот говорит (или, в случае дельфинов, свистит)26 . И у китообразных, и у певчих птиц, как и у человека, вокализация произвольна. Она независима от лимбической системы (подкорковых структур), не свидетельствует об эмоциональном возбуждении и осуществляется скелетной мускулатурой27 . Органы же звукопроизводства при этом совершенно разные: у человека это прежде всего гортань с голосовыми связками, у дельфинов и китов – назальные мешки, у птиц – сиринкс (иначе “нижняя гортань”, расположенная не в начале трахеи, как гортань млекопитающих, а в том месте, где от трахеи ответвляются бронхи; эволюционное происхождение сиринкса и гортани млекопитающих различно).

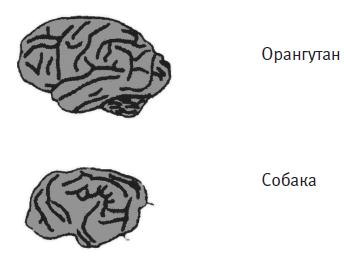
Рис. 4.4. Мозг дельфина, человека, орангутана и собаки.
У китообразных, как и у певчих птиц, имеется латерали‑зация мозга. Но если у китообразных, как и у человека, асимметрично устроена кора больших полушарий (неокортекс), то у птиц это свойство реализовано на базе хотя и гомологичных новой коре, но все же не тождественных ей структур – нидопаллиума и гиперпаллиума (раньше их называли неостриатум и гиперстриатум соответственно)28 .
Впрочем, асимметрия мозговых структур обнаруживается у самых разных животных, в том числе у угрей, тритонов, лягушек и акул29 .
И для китообразных, и для певчих птиц чрезвычайно важна способность к звукоподражанию. Так, дельфины заимствуют “свист‑подпись” у других дельфинов той же группы. Впрочем, способность к звукоподражанию была обнаружена у целого ряда видов, пользующихся звуковой коммуникацией, – она есть не только у певчих птиц и у китообразных, но и у летучих мышей, тюленей30 , слонов31 , а возможно, даже у мышей. Способность к обучению звуковым элементам коммуникации, по‑видимому, характерна прежде всего для тех видов, у которых звук используется для поддержания социальной структуры.
Все эти (и другие, которые наверняка будут еще открыты) сходные черты коммуникативных систем певчих птиц, китообразных и человека, как можно видеть, приобретены независимо. Поскольку эти сходства охватывают целый комплекс свойств, их возникновение в ходе эволюции, вероятно, представляло собой процесс с положительной обратной связью, и ответ на вопрос о том, что является причиной, а что следствием, далеко не очевиден. В частности, по мнению Т. Дикона, асимметрия, присущая человеческому мозгу, является скорее следствием, нежели причиной возникновения языка32 .
Изучение коммуникации животных позволяет разрешить самую непостижимую для некоторых исследователей “загадку языка” – почему он вообще возможен. Действительно, особь, производящая коммуникативные действия, тратит свое время и силы, становится более заметной хищникам – ради чего? Зачем передавать информацию другим вместо того, чтобы воспользоваться ею самому33 ? Почему бы не обмануть сородичей, чтобы получить свою выгоду34 ? Зачем пользоваться информацией от других, а не собственными ощущениями35 ? Или, может быть, выгоднее собирать информацию на основе сигналов других особей, а самому “молчать” (тем самым не платя высокую цену за производство сигнала)? Подобные рассуждения приводят, например, к идее, что язык развился для манипулирования сородичами (см. подробнее ниже, гл. 5). Или, может быть, появление языка вообще не связано с информационным обменом? Может быть, язык возник исключительно как инструмент мышления, как считает Ноам Хомский, или даже вообще в качестве игры, как полагает антрополог Крис Найт36 ?
В самом деле, если анализировать действие естественного отбора на индивидуальном, а не на групповом уровне, то преимущества коммуникативной системы (любой – не только языка) обнаружить не удается. И это приводит некоторых исследователей к выводу, что естественный отбор не играл никакой роли в процессе глоттогенеза37 , и возникновение языка, возможно, в принципе не связано с обретением каких‑либо адаптивных преимуществ, а просто является побочным эффектом развития каких‑то других свойств, например, прямохождения (см. гл. 3)38 .
Но на самом деле все перечисленные выше вопросы могут быть отнесены не только к человеческому языку – они релевантны для любой коммуникативной системы. И задавать их может лишь человек, не искушенный в этологии. Действительно, любая коммуникация – дело затратное: животное расходует энергию на производство сигнала, тратит время (которое могло бы быть использовано для чего‑то, приносящего непосредственную биологическую пользу, например, для питания или гигиенических процедур), во время производства и восприятия сигнала менее внимательно следит за всем остальным, рискуя быть съеденным (классический пример – токующий глухарь, см. фото 19 на вклейке). Кроме того, энергия тратится на поддержание мозговых структур, необходимых для восприятия сигналов, и анатомических структур, необходимых для их производства. Однако “альтруистическое” поведение коммуницирующих особей, идущих на определенные затраты ради того, чтобы (вольно или невольно) передать своим сородичам информацию, ведет в итоге к общему увеличению количества “альтруистов” – даже если внутри своей популяции они проигрывают конкурентную борьбу более “эгоистичным” сородичам, – поскольку популяции, в которых альтруистов много, увеличивают свою численность гораздо более эффективно, чем популяции с преобладанием “эгоистов”. Этот статистический парадокс, известный как “парадокс Симпсона” {29}, был недавно смоделирован на бактериях39 , среди которых также есть особи, отличающиеся “альтруистическим” поведением, т. е. производящие – с повышением собственных затрат – вещества, способствующие росту всех окружающих бактерий. Чем сильнее конкуренция между группами, тем выше оказывается уровень альтруизма и кооперации внутри отдельных групп40 .
Коммуникативная система – любая – возникает, развивается и существует не для выгоды особи, подающей сигнал, и не для выгоды особи, его принимающей; ее назначение – даже не организация отношений в паре “говорящий” – “слушающий”. Коммуникативная система представляет собой “специализированный механизм управления в системе популяции в целом”41 .
Особи одного вида неизбежно оказываются конкурентами друг друга, поскольку претендуют на одни и те же ресурсы (пищу, укрытия, половых партнеров и т. д.). Тем не менее, при выборе места обитания животные предпочитают селиться по соседству с представителями своего вида. Соседство может быть тесным (как, например, у групповых млекопитающих или колониальных птиц) или не очень (например, индивидуальные участки тигров или медведей простираются на многие километры), но даже медведи не стремятся поселиться там, где бы поблизости вообще не было других медведей. И понятно почему: если бы появилась особь, в генах которой было бы заложено стремление поселиться как можно дальше от сородичей (и тем самым избавиться от конкурентов), ей было бы крайне трудно найти себе пару и передать эти гены потомству. Как показали недавние исследования42 , птицы выбирают гнездовые участки рядом с участками сородичей, но стремятся селиться подальше от представителей видов, занимающих сходную экологическую нишу. Это значит, что конкуренция за ресурсы между представителями одного вида и разных видов устроена неодинаково: если чужаков лучше избегать или выгонять, то со своими можно “договориться” – при помощи коммуникативных взаимодействий распределить ресурсы так, чтобы этих ресурсов (пусть и разного качества) в итоге хватило всем.
Коммуникативная система позволяет каждой особи находить свое место. Например, особь, получившая по итогам коммуникативных взаимодействий высокий ранг, может кормиться тем, что дает много энергии, но требует больших временных затрат на то, чтобы подготовиться к добыче корма самым специализированным и эффективным методом, – она “знает”, что ее не будут беспокоить слишком часто. Особь же низкоранговая выберет такую пищедобывательную стратегию, которая не сулит большой энергетической выгоды, но зато позволяет часто отвлекаться. И это дает существенный выигрыш, поскольку попытка добывать высокопитательный, но затратный по времени корм обернулась бы для низкоранговой особи настоящей трагедией: среди ее соседей слишком много охотников “самоутвердиться за ее счет” (т. е. повысить свой ранг за счет коммуникативной победы над ней), и реализовать такую стратегию кормления она бы просто не успела. Таким образом, коммуникация значительно ослабляет конкуренцию за ресурсы и позволяет выжить большему количеству представителей одного вида. Подобным же образом коммуникация распределяет особей и в других важных для жизни вида аспектах, например, при половом размножении. Так, высокоранговый олень завоевывает себе целый гарем самок и получает возможность передать свои гены большому количеству потомков. А низкоранговые олени, не имеющие собственного гарема, получают доступ к противоположному полу иначе: потихоньку, пока хозяин гарема не видит, они спариваются с его самками и тем самым тоже обеспечивают себе определенный репродуктивный успех43 .
Кроме того, у видов, практикующих половое размножение, имеется задача “морально подготовить” партнеров к спариванию. Решение такого рода задач без посредства коммуникативной системы воистину “смерти подобно” – это наглядно показывают австралийские сумчатые мыши (род Antechinus ). Их самцы кидаются на самок, “не говоря ни слова” (т. е. без предварительного обмена какими‑либо коммуникативными сигналами), – и в итоге ни один из них не переживает сезона размножения. Как показали данные Иэна Мак‑Дональда и его коллег44 , все погибают от стресса, хотя в принципе организм самца сумчатой мыши рассчитан на более долгую жизнь: если держать его дома в клетке, не подпуская к самкам (и другим самцам, с которыми он также вступал бы в физические, а не в коммуникативные взаимодействия), он проживет примерно года два, как и самка.

Рис. 4.5. Сумчатая мышь – живое доказательство того, что без коммуникации жить можно, но плохо и недолго.
При высокой плодовитости и отсутствии эффективных хищников такой вид еще может существовать, но при менее благоприятных условиях он, вероятно, не выдержал бы конкуренцию с видами, пользующимися коммуникацией.
Наличие в репертуаре вида специальных коммуникативных действий позволяет уменьшить количество прямых физических воздействий на сородичей: если особи могут, обменявшись несколькими сигналами, выяснить, кто из них выше другого в иерархии, имеет больше прав на самку и т. д., отпадает нужда кусать, клевать или как‑либо иначе травмировать друг друга. Соответственно, чем более совершенна коммуникативная система вида, тем менее опасными для здоровья партнеров оказываются процессы взаимодействия.
Развитая коммуникативная система дает возможность эффективно организовывать совместную деятельность нескольких особей – даже если в процессе этой деятельности сигналы и не используются. Так, например, волки, которые не имели случая ранее “договориться” между собой о взаимной иерархии, не могут слаженно охотиться на оленя (и, соответственно, вынуждены довольствоваться полевками и другими грызунами). Непосредственно в момент охоты волки не обмениваются сигналами, но “понимание” своего места в иерархии задает некий внутренний ритм движений каждого животного. Совокупность дополняющих друг друга различных “внутренних ритмов” позволяет успешно объединять усилия45 .
Еще одна задача коммуникативной системы – сортировка особей по территориям. Те, кто коммуницирует успешнее других, имеют наибольшие шансы занять максимально удобные местообитания {30} (т. е. такие, к которым особи данного вида лучше всего приспособлены). Менее успешные коммуниканты оттесняются на периферию. Таким образом коммуникативная система организует структуру популяции, и это позволяет – не конкретным особям, а именно популяции в целом – формировать приспособительный ответ на изменения экологической ситуации.
В целом можно сказать, что возможность общаться позволяет виду (прежде всего именно виду, а не отдельным его представителям) сдвигать свою активность с непосредственной реакции на уже происшедшие события в область экстраполяции и прогноза46 : в результате действий, которые совершаются не “в пожарном порядке” (после того, как нечто случилось), а в относительно комфортных условиях готовности к общению, будущее оказывается до какой‑то степени доступным прогнозированию. Обмен сигналами позволяет особи составить некоторый прогноз на будущее – и действовать, исходя из него. Соответственно, преимущество получают те особи, которые умеют строить свою активность при условии знания , что их ждет дальше. Это обеспечивает виду бóльшую стабильность. Чем более совершенна коммуникативная система, тем в большей степени будущее в результате ее применения становится предсказуемым (а впоследствии и формируемым). Кроме того, “коммуникативная система стимулирует развитие самых разных компенсаторных механизмов у всех, говорящих “не так””47 , поскольку “общение продолжается даже при нарушениях в правилах передачи знаков, если партнеры готовы менять установки в сторону нормы ”48 .
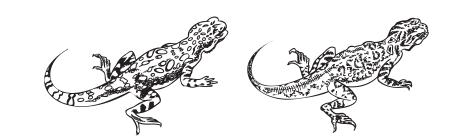
Рис. 4.6. Такырная круглоголовка (слева) лучше вооружена, чем ее близкая родственница – сетчатая круглоголовка (справа). Поэтому для такырной круглоголовки оказывается полезным использовать коммуникативные сигналы вместо прямых физических воздействий. А для сетчатой круглоголовки, наоборот, выгоднее «сэкономить» на коммуникации: поскольку ее укусы не так страшны, тратить много ресурсов на избавление от них нерентабельно.
Как возникают коммуникативные сигналы, можно наблюдать на примере двух близких видов ящериц – такырной и сетчатой круглоголовок (Phrynocephalus helioscopus, Ph. reticulatus )49 . Для круглоголовок необходимо, чтобы самец не спаривался с самкой, которая уже оплодотворена другим самцом (и не тратил попусту свои репродуктивные ресурсы). Соответственно, самка должна уклониться от спаривания. Сетчатая круглоголовка в таких случаях либо убегает, либо кусает самца. Но у такырных круглоголовок такой номер не пройдет: во‑первых, такырные круглоголовки более целеустремленные, значит, тактика “убежать” потребует бóльших затрат. А во‑вторых, они лучше вооружены, так что укусы нанесут более серьезный ущерб здоровью самца. И тогда возникает коммуникативный сигнал. Легко заметно, что это, в сущности, те же самые движения, что и у сетчатой круглоголовки: движения, отражающие конфликт двух побуждений – убежать и укусить. Но если у сетчатой круглоголовки эти движения определяются чисто эмоционально и могут быть вообще незаметны, то такырная круглоголовка делает их явно напоказ: они более стереотипны, даже несколько неестественны, с резкими, четко выделимыми границами, вся демонстрация продолжается дольше, чем у сетчатой круглоголовки. И это неудивительно: для такырных круглоголовок очень важно, чтобы самец отказался от своих намерений без ущерба для здоровья – как своего, так и самки.
Заметим, что о сколь‑нибудь настоящей “сигнализации” тут речь, возможно, и не идет. Самка не хочет ничего сообщить самцу, она просто испытывает очень сильные колебания между намерением укусить и намерением убежать – настолько сильные, что самец успевает заметить этот конфликт мотиваций, и у него запускается – опять‑таки, без всякого, вероятно, участия сознания – поведение “прекратить преследование”. И отбор благоприятствует тем популяциям, где чаще рождаются самки, способные максимально тщательно продемонстрировать самцу свои намерения, и самцы, с максимальной эффективностью распознающие демонстрацию самки. Соответственно, у самцов формируются детекторы для обнаружения характеристических черт самочьей “пантомимы”, а самки делают свои движения все более четкими и стереотипными, такими, чтобы их явно очерченные границы максимально хорошо распознавались детекторами самца. Кроме того, демонстрация самки продолжается заметное время – с тем чтобы самец успел распознать сигнал и запустить соответствующую программу поведения.
Впрочем, справедливости ради следует отметить, что у такырных круглоголовок (как, впрочем, и у нас, людей) случаются “коммуникативные неудачи”, так что некоторые самцы в итоге становятся жертвой укусов. Но доля таких самцов существенно (статистически значимо) меньше, чем у сетчатой круглоголовки.
Этот пример наглядно показывает, что для возникновения коммуникативных сигналов не нужен гений, в порыве вдохновения творящий знаки, изобретающий всё новые сочетания форм и смыслов. Не нужно, вероятно, даже сознание. Необходимо лишь, чтобы нервная система могла отслеживать события, происходящие во внешнем мире, и запускать оптимально отвечающие им поведенческие программы. Если для жизни вида окажется важным, чтобы о тех или иных намерениях особи ее сородичи могли узнавать до того, как эти намерения воплотятся в действия, отбор позаботится о том, чтобы сделать соответствующие намерения максимально заметными – с одной стороны, акцентировать некоторые компоненты физических проявлений соответствующего намерения, а с другой – настроить детекторы на их распознавание. Стандартный путь развития коммуникативных систем состоит в том, что особи наблюдают за внешним видом и/или поведением сородичей и у них формируются детекторы для регистрации этого. Вместе с тем элементы внешнего вида и/или поведения сородичей становятся всё более легко регистрируемыми при помощи детекторов. Возникает положительная обратная связь между отправителем и получателем коммуникативного сигнала, заставляющая коммуникативную систему все более – в эволюционной перспективе – усложняться (разумеется, лишь до тех пор, пока затраты на коммуникацию не начнут превышать выгоды от нее). Создать детекторы, регистрирующие те или иные характеристики сородичей, эволюционно проще, чем создать детекторы, пригодные для наблюдения за другими видами, ландшафтом и т. п. (хотя и такие детекторы, разумеется, тоже имеются у организмов), поскольку и бóльшая заметность элементов внешнего вида и/или поведения, и степень восприятия их кодируются в одном и том же геноме и подвергаются фактически одному и тому же естественному отбору.
В принципе, любое поведение животного его сородичи могут заметить и изменить в связи с этим свое собственное поведение. Например, когда голубь клюет ломоть хлеба, другой голубь (или, скажем, воробей) может, увидев это, приблизиться и начать клевать тот же ломоть с другого конца (если, конечно, его не прогонят). Поэтому в животном мире нередки действия, которые имеют как информационную, так и неинформационную составляющую. Например, такими являются действия собаки, метящей территорию собственной мочой: для того, чтобы опорожнить мочевой пузырь, ей достаточно было бы помочиться однократно (а не поднимать лапку у каждого дерева или столба, роняя всякий раз по нескольку капель), но оставленный запах несет информацию для других собак.
О собственно “сигналах”, возможно, следует говорить лишь тогда, когда то или иное действие перестает приносить непосредственную биологическую пользу, становясь только средством передачи информации. В этом случае оно оптимизируется не под изменчивые характеристики окружающего мира, а под жестко настроенные детекторы.
Возможно, именно в грубой работе детекторов разгадка того, почему движения, перешедшие из области обычной повседневной активности в сферу коммуникации, часто становятся резкими и “вычурными”, а их отдельные элементы выдерживаются дольше, чем сходные элементы обычного поведения. Например, райские птицы, демонстрируя, могут часами висеть вниз головой.
Такого рода дискретные, длительно выдерживаемые сигналы зафиксированы у птиц и рептилий, у млекопитающих же во многих случаях структура коммуникативной системы иная. Может быть, дело в том, что кора больших полушарий головного мозга (неокортекс) дает возможность более эффективного распознавания, может быть, в чем‑то еще, но у млекопитающих коммуникативные сигналы часто оказываются континуальными, с бесконечным количеством переходных ступеней от одного сигнала к другому. На рисунке 4.7 изображена мимика домашней кошки, соответствующая разным степеням страха и агрессивности. На схеме показаны лишь по три градации для каждой из эмоций, но, разумеется, кошка не автомат, который резко “перещелкивается” из позиции 1 в позицию 2 и далее в позицию 3. Читатель может сам мысленно достроить то бесконечное количество оттенков обоих этих чувств, которое займет промежуточную позицию между любыми двумя соседними клетками данной схемы.
Впрочем, у млекопитающих есть не только эмоциональные сигналы, плавно переходящие один в другой. Сравнительное изучение разных видов, относящихся к одной классификационной группе (т. е. к одному таксону), дает возможность увидеть тенденции развития коммуникативных систем.
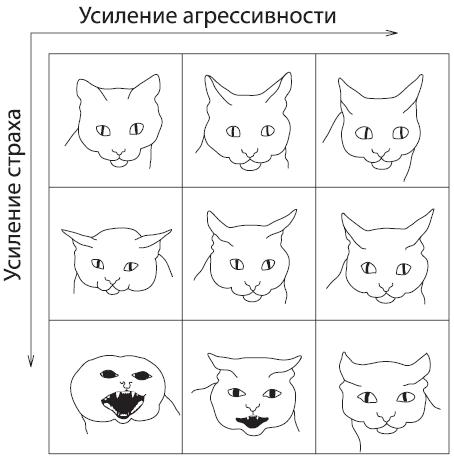
Рис. 4.7. Мимика домашней кошки 50 .
Рассмотрим в качестве примера два разных вида сусликов (см. фото 20 на вклейке) – более примитивного (по своему строению) калифорнийского суслика (Spermophilus beecheyi ) и более “прогрессивного” суслика Белдинга (Spermophilus beldingi ). У обоих видов есть сигналы опасности – щебет и свист. У суслика Белдинга свист – сигнал очень сильной опасности, а щебет (или, точнее, его аналог – трель) – умеренной. Заметим еще раз, что под словом “сигнал” здесь не имеется в виду никакого намеренного действия, специально предназначенного для коммуникации. Просто у суслика, который сильнее испуган, звук получается более похожим на свист – тем более, чем сильнее страх. Соответственно, между трелью и свистом возможно бесконечное число промежуточных “сигналов”. Сородичи, слышащие этот звук, “заражаются” соответствующей эмоцией (подобно тому, как людей “заражает” зевота или смех), и у многих из них непроизвольно возникает соответствующая вокализация. К этому уровню развития коммуникации вполне применимо рассуждение Е.Н. Панова51 , согласно которому никаких “языков” у животных нет.
Но у калифорнийского суслика коммуникативная система устроена принципиально иначе. Свист и щебет становятся референциальными сигналами (англ. referential signals ), т. е. сигналами, обозначающими вполне конкретный объект внешнего мира (называемый в семиотике “референтом”): свист означает “опасность с воздуха”, щебет – “опасность с земли”52 .
“Этимология” этих сигналов не менее прозрачна, чем “этимология” демонстраций такырной круглоголовки: летящий хищник обычно более опасен (и, соответственно, страшен), чем хищник наземный. Но функционирование свиста и щебета у калифорнийского суслика отличается кардинально. Промежуточных градаций между ними нет – как нет промежуточных градаций между орлом, летящим по воздуху, и койотом, бегущим по земле. Эти сигналы уже не настолько связаны с эмоциями: суслик может быть очень испуган внезапным появлением наземного хищника, но все равно звук, который он издаст, будет (с максимальной вероятностью) щебетом, а не свистом. И наоборот, хищная птица может быть очень далеко в небе и не вызывать большого страха – но суслик, видя ее, будет (в подавляющем большинстве случаев) издавать свист. Сигналы этого типа (хотя они, возможно, также не являются преднамеренными) не “заражают” сородичей эмоциями, а предоставляют им конкретную информацию об окружающем мире.
Соответственно, референциальные сигналы с полным правом можно назвать сигналами‑символами (как это сделано в работе этолога Владимира Семеновича Фридмана53 ), поскольку у них нет обязательной природной связи между формой и смыслом. Интересно, что у этих видов сусликов различается и восприятие сигнала: суслики Белдинга ретранслируют сигнал только в том случае, если сами в достаточной степени напуганы, калифорнийские же суслики способны передавать информацию дальше независимо от своего эмоционального состояния. Интенсивность воздействия сигнала в этой системе пропорциональна не степени возбуждения издающей сигнал особи, а степени стереотипности его внешней формы (поскольку наиболее “правильного” вида сигналы наиболее эффективно распознаются детекторами).
Этот пример показывает, что специализация к определенному типу существования у общественных животных может предполагать не только те или иные анатомические изменения, но и оптимизацию “заметных” действий (коммуникативных сигналов), их освобождение от эмоций и обретение ими способности обозначать конкретные объекты (или ситуации) окружающего мира. Именно на этом уровне развития коммуникативной системы возникает не только произвольность знака, но и возможность оторваться от “здесь и сейчас”: суслику достаточно услышать свист, чтобы мочь запустить поведенческий комплекс, обеспечивающий спасение от хищной птицы, – наблюдать самого хищника ему при этом необязательно. Отрыв от “здесь и сейчас” позволяет особи принимать менее эмоциональное, более “взвешенное” решение о том, что следует делать дальше.
Референциальные сигналы, подобно элементам человеческого языка, характеризуются категориальным восприятием. Это было проверено, в частности, в опытах Алексея Анатольевича Шибкова на самых примитивных представителях отряда приматов – тупайях (Tupaia glis , см. фото 21 на вклейке). Совмещая подачу одного из сигналов, присущих данному виду, со слабым ударом электрическим током, у животных вырабатывали вполне заметную реакцию на данный сигнал – реакцию избегания. Потом характеристики сигнала плавно меняли, постепенно превращая его в другой сигнал того же самого вида. В полном соответствии с моделью категориального восприятия, пока сигнал оставался “тем же самым” (по мнению подопытной тупайи), животные демонстрировали реакцию избегания, но как только сигнал становился “другим”, эта реакция немедленно исчезала54 .
Системы референциальных сигналов были обнаружены у многих видов животных – у сурикатов (африканских мангустов) Suricata suricatta (различаются типы опасности – наземный хищник, хищная птица, змея)55 , у кошачьих лемуров Lemur catta (различаются “опасность с земли” и “опасность с воздуха”)56 , у луговых собачек (наземных грызунов из семейства беличьих) Cynomys gunnisoni 57 и даже у домашних кур (обозначение двух типов опасности – наземный и воздушный хищники – и “пищевой” крик)58 . Вероятно, развитие таких сигналов из эмоциональных является эволюционной тенденцией – оно прослеживается, в частности, у сурков59 .
Из референциальных сигналов состоит система предупреждения об опасности у верветок (Cercopithecus aethiops , см. фото 22 на вклейке). Как установили приматологи Дороти Чини и Роберт Сифард60 , у верветок имеются четко различающиеся сигналы опасности: один крик обозначает орла, другой – леопарда (или гепарда), третий – змею (мамбу или питона), четвертый – опасного примата (павиана или человека). Исследователи проигрывали им магнитофонные записи разных типов криков (в отсутствие соответствующих опасностей), и верветки всякий раз реагировали “правильно”: по сигналу “леопард” бросались на тонкие верхние ветки, по сигналу “орел” спускались на землю, по сигналу “змея” вставали на задние лапы и осматривались. Чтобы выяснить, являются ли сигналы верветок эмоциональными или референциальными, исследователи делали записи длиннее или короче, громче или тише – для эмоциональных сигналов именно эти характеристики имеют основное значение, для референциальных же они совершенно несущественны (подобно тому, как для смысла слова в общем случае не важно, будет ли оно произнесено быстро или медленно, громко или тихо). Опыты показали, что для верветок важна не интенсивность сигнала, а его формантные характеристики.
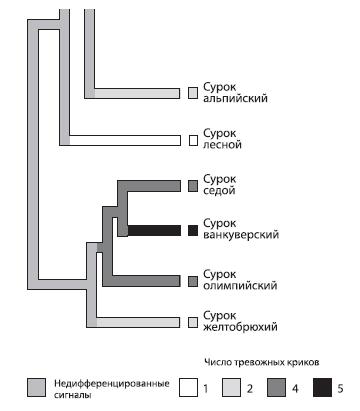
Рис. 4.8. Это родословное древо сурков (род Marmotta) построено на основании молекулярных данных, но по нему видно, что при переходе от более примитивных видов к более прогрессивным число различных сигналов увеличивается 61 .
Коммуникативную систему верветок нередко рассматривают как промежуточный этап на пути к человеческому языку: сначала сигналов было, как у верветок, лишь несколько, потом, постепенно добавляя по одному сигналу, предки человека добрались в конце концов до языка современного типа62 . Однако это, по‑видимому, неверно. Дело в том, что, во‑первых, внешняя форма (звуковая оболочка) сигналов у верветок является врожденной, следовательно, расширение такой коммуникативной системы и добавление в нее новых сигналов может происходить только путем генетических мутаций. Человеческая же система знаков не врожденная, она содержит огромное число элементов (десятки тысяч – для такого количества нужных мутаций просто не хватило бы эволюционного времени) и, кроме того, является принципиально открытой, добавление в нее новых знаков легко происходит за время жизни одного индивида. Возможно, что вы, читая эту главу, пополнили свой лексикон несколькими новыми словами – верветке такого не достичь. Все, что она может сделать за время жизни, – это несколько уточнить форму (акустические характеристики) и значение того или иного крика (например, усвоить, что сигнал “орел” не относится к птицам‑падальщикам) {31}.
Во‑вторых, в человеческом языке принципиально иначе устроена реакция на сигнал. Если у верветок восприятие сигнала жестко задает поведение, то у человека восприятие сигнала задает лишь начало деятельности по его интерпретации (по мнению Т. Дикона, это вызвано наличием огромного количества ассоциативных связей между словами‑символами в мозге64 ), результаты же этой интерпретации могут зависеть от личного опыта, от индивидуальных особенностей характера, от отношения к подавшему сигнал, от сиюминутных намерений и предпочтений и т. д. и т. п. Поэтому нередко оказывается, что реакция на один и тот же текст у разных слушателей (или читателей) резко различается.
Подобная разница между людьми и верветками вполне объяснима. У верветок функция данного фрагмента коммуникативной системы состоит в том, чтобы обеспечить быстрый запуск правильной поведенческой программы спасения от соответствующего хищника, поэтому любые отклонения от стандартной реакции подавляются отбором. Человек же, в значительной степени вышедший из‑под контроля естественного отбора, может себе позволить долгие размышления о смысле услышанного сообщения. Таким образом, хотя верветки относятся, как и мы, к отряду приматов, между их коммуникативной системой и языком нет гомологии, а есть лишь аналогия.
У других представителей церкопитековых, больших белоносых мартышек (Cercopithecus nicticans , см. фото 23 на вклейке), можно наблюдать еще одну аналогию с человеческим языком65 . У этих мартышек, как и у верветок, есть разные сигналы для разных типов опасностей – крик “пяу” (в англоязычных работах – pyow ) означает “леопард”, крик “хак” (hack ) – “орел”. Но у них, как установили Кейт Арнольд и Клаус Цубербюлер, есть также возможность комбинирования сигналов, и при этом получается, как в человеческом языке, нетривиальное приращение смысла (не сводящееся к простой сумме смыслов составных частей). Когда самец произносит последовательность “пяу‑хак” (или, чаще, повторяет каждый из этих криков несколько раз – но именно в такой последовательности), это вызывает не реакцию спасения от леопарда или орла, а перемещение всей группы на достаточно значительное расстояние – более значительное, чем без сигнала “пяу‑хак”. Некоторые исследователи склонны видеть в этом сходство с человеческим синтаксисом (два “слова” составляют “предложение”), другие полагают, что это больше напоминает морфологию (сложное слово типа кресло ‑качалка ), но это не более чем спор об аналогии. В качестве же гомологии с языком здесь можно рассматривать лишь когнитивную возможность получать при комбинировании сигналов нетривиальное приращение значения (ср. вечер – вечерник “студент вечернего отделения института”, но утро – утренник “праздник или представление, устраиваемое утром”: один и тот же суффикс в сочетании с названиями разных частей дня добавляет совершенно разный смысл).
Еще более развернутую аналогию с человеческим языком можно усмотреть в коммуникативной системе мартышек Кемпбелла (Cercopithecus campbelli campbelli , см. фото 24 на вклейке), живущих в национальном парке Таи (Берег Слоновой Кости). Самцы этих обезьян употребляют шесть видов сигналов, которые исследователи (К. Цубербюлер и его соавторы) записывают как “бум”, “крак”, “крак‑у”, “хок”, “хок‑у” и “вак‑у”66 . Элемент “‑у”, выделяемый в трех из этих сигналов, авторы интерпретируют как суффикс. Он, подобно, например, русскому суффиксу – ств (о ) (ср. братство ) или английскому – hood (ср. brotherhood “братство” от brother “брат”), не употребляется отдельно, но определенным образом изменяет значение той основы, к которой присоединяется. Так, сигнал “крак” обозначает леопарда, а сигнал “крак‑у” – опасность вообще.
Комбинирование знаков дает, как и у больших белоносых мартышек, нетривиальные приращения смысла. Например, серия криков “крак‑у” может быть издана, когда мартышка слышит голос леопарда или крик мартышек диан, предупреждающий о появлении леопарда, но если этому сигналу предшествует дважды повторенный сигнал “бум”, то вся “фраза” интерпретируется как “падает дерево или большая ветка”. Если в серию криков “крак‑у”, которой предшествует пара криков “бум”, иногда вставлять крик “хок‑у”, получится территориальный сигнал, который самцы издают при встрече на границе участка с другой группой мартышек Кемпбелла. Просто двукратное повторение крика “бум” означает, что самец потерял из виду свою группу (самки, слыша такой сигнал, подходят к самцу). Всего авторы выделили девять возможных “фраз”, скомбинированных из этих шести криков.
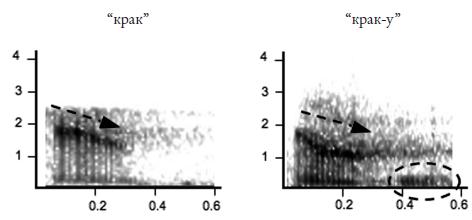
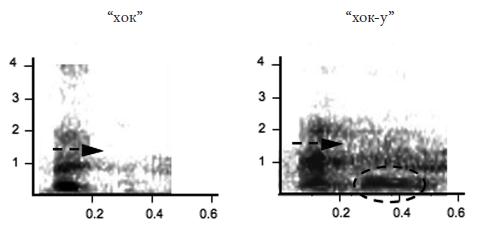
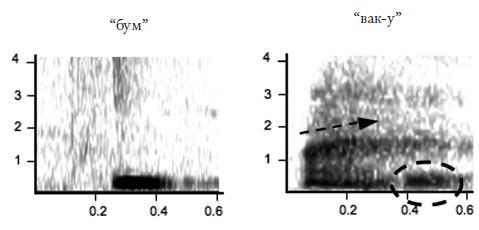
Рис. 4.9. Звуковые сигналы мартышек Кемпбелла (сонограммы). Черная стрелка показывает движение формант; пунктирной рамкой обведен “суффикс” “‑у” 67 .
В коммуникативной системе мартышек Кемпбелла представлены и правила “порядка слов”: например, сигнал “бум” употребляется только в начале цепочки криков и всегда повторяется дважды, сигнал “хок” предшествует сигналу “хок‑у”, если они встречаются вместе, серия криков, предупреждающая об орле, начинается обычно с нескольких криков “хок”, а заканчивается несколькими криками “крак‑у” и т. д.
По мнению авторов исследования, в некоторых аспектах эта коммуникативная система приближается к человеческому языку даже больше, чем успехи человекообразных обезьян, обученных языкам‑посредникам и умеющих составлять комбинации типа “ВОДА”+“ПТИЦА”, хотя настоящей грамматикой она все же не обладает68 . И дело здесь не только в том, что правила достаточно просты, а их число невелико. Главное, на мой взгляд, отличие этой системы от человеческого языка – отсутствие в ней достраиваемости: есть шесть криков и девять возможных “предложений”, и этим все ограничивается, новые знаки и новые сообщения не строятся.
Ограниченность исследованного материала не дает возможности судить о том, являются ли все эти сигналы (в том числе содержащие суффикс “‑у”) и их комбинации врожденными, присущими всем представителям Cercopithecus campbelli campbelli , или же, по крайней мере, некоторая часть этой системы является культурной традицией данной конкретной популяции. По наблюдениям авторов, верно скорее первое: сигналы издаются без волевого контроля, самцы не демонстрируют намерения информировать сородичей, они просто испытывают эмоции – и на этом фоне у них издаются соответствующие крики. В то же время эти данные показывают, что даже при отсутствии волевого контроля за звукопроизводством жизнь вида, ведущего групповой образ жизни, в лесу, в условиях низкой видимости и большого количества хищников располагает к формированию коммуникативной системы, которая использует комбинации звуковых сигналов (как друг с другом, так и с элементами, не являющимися отдельными сигналами), чтобы из небольшого количества доступных врожденных криков произвести больше различных сообщений.
Если рассмотреть коммуникативные системы различных видов позвоночных, можно увидеть еще одну общую тенденцию – уменьшение степени врожденности. У низших животных, обладающих коммуникативной системой, врожденной является как внешняя форма сигнала, так и его “смысл” (то, что так или иначе будет определять поведение животного, воспринявшего данный сигнал); реакция на сигнал так же врожденна и стереотипна, как и реакция на несигнальные раздражители (поэтому такие сигналы называются релизерными). Например, птенец серебристой чайки, выпрашивая пищу, клюет красное пятно на клюве родителя, и это побуждает родителя покормить птенца, – в этом примере врожденными, инстинктивными, являются как действия птенца, так и реакция взрослой птицы. Сигналы такого рода, разумеется, могут до некоторой степени совершенствоваться в ходе развития отдельной особи (так, птенец чайки с течением времени “натренировывается” более метко попадать в красное пятно), но не более, чем любые другие инстинктивные действия.
У животных, обладающих более высоким уровнем когнитивного развития, появляются так называемые “иерархические” сигналы. Этот термин, введенный этологом В.С. Фридманом, подчеркивает, что основная функция этих сигналов – обслуживание иерархических отношений между особями в пределах группировки. Форма иерархических сигналов еще является врожденной, но “смысл” устанавливается в каждой группировке отдельно. Например, предъявление большим пестрым дятлом своему сородичу крайних рулевых перьев обозначает “это я”, смысл же “эта особь выше меня по иерархии” (или “эта особь ниже меня по иерархии”) сородич, увидевший этот сигнал, достраивает, исходя из опыта предыдущих взаимодействий с данной птицей. Такой смысл не может быть врожденным, поскольку невозможно предугадать заранее место конкретной особи в конкретной группировке. Кроме того, такой смысл может меняться по итогам взаимодействия особей друг с другом.
Следующая ступень развития – так называемые “ad‑hoc‑сигналы”, имеющиеся лишь у узконосых обезьян (начиная с павианов): эти элементы коммуникативного поведения создаются по ходу дела, для сиюминутных нужд, соответственно, врожденными не являются ни их форма, ни их “смысл”. Такую коммуникативную систему может себе позволить лишь вид с достаточно хорошо развитым мозгом, поскольку, чтобы поддерживать коммуникацию такого рода, особи должны быть готовы придавать сигнальное значение действиям, до этого сигналами не являвшимся.
Человеческий язык представляет собой следующий член этого ряда: бывшие ad‑hoc‑сигналы начинают закрепляться, накапливаться и передаваться по наследству посредством обучения и подражания – так же, как, например, умение изготавливать орудия труда. В результате получается “инструментальная” (термин А.Н. Барулина) семиотическая система.
В качестве одного из наиболее существенных отличий коммуникативных систем животных от человеческого языка нередко называется то, что они не связаны с индивидуальным опытом, с рассудочной деятельностью, тогда как у человека язык и мышление объединились в ходе эволюции “в одну речемыслительную систему”69 . Действительно, сигналы с врожденной формой и врожденным смыслом не могут передавать жизненный опыт отдельной особи – только обобщенный опыт вида. Но уже иерархические сигналы отчасти отражают индивидуальный опыт, хотя и лишь в одной, весьма ограниченной области, – опыт конкурентных взаимодействий одной особи с другими. Еще в большей степени связаны с личным опытом ad‑hoc‑сигналы, поскольку в них как форма, так и смысл могут включать то, что стало известно конкретной особи в течение ее жизни (см. ниже).
Что же касается обезьян, то их звуковые сигналы, хотя и являются по форме врожденными, также, вероятно, могут участвовать в передаче личного опыта. Свидетельницей одного такого случая стала С. Сэвидж‑Рамбо после вечерней прогулки по лесу с бонобо Панбанишей. Во время прогулки они заметили на дереве силуэт какой‑то крупной кошки и, испугавшись, вернулись в лабораторию, где их встретили бонобо Канзи, Тамули, Матата и шимпанзе Панзи. Обезьяны (вероятно, по невербальным сигналам) догадались, что Панбанишу и С. Сэвидж‑Рамбо что‑то напугало в лесу – они, пишет Сэвидж‑Рамбо, “стали напряженно всматриваться в темноту и издавать мягкие звуки “уху‑ух”, говорящие о чем‑то необычном. <Панбаниша> тоже начала издавать какие‑то звуки, как будто рассказывала им о большой кошке, которую мы видели в лесу. Все остальные слушали и отвечали громкими криками. Неужели она говорит им что‑то, чего я не могу понять? Я не знаю”70 . Какую информацию передала Панбаниша, в точности сказать трудно (йеркишем она не воспользовалась), но “Канзи и Панзи, когда им в очередной раз разрешили погулять, обнаружили колебания и страх именно в этом участке леса. Поскольку их прежде никогда не пугали, похоже, все‑таки, что они смогли что‑то понять из происшедшего”71 .
Подобный же “рассказ” наблюдала и отечественный приматолог Светлана Леонидовна Новоселова. Шимпанзе Лада, которую однажды пришлось, несмотря на ее отчаянный вой и сопротивление, вынести на прогулку, на следующий день “поведала” людям о случившемся: “Обезьяна, драматически воздев руки, привстала в своем гнезде на широкой полке, спустилась и, бегая по клетке, воспроизвела интонационно очень верно в своем крике, который продолжался не менее 30 мин, эмоциональную динамику переживаний предшествующего дня. У меня и у всех окружающих сложилось полное впечатление “рассказа о пережитом””72 .
Такое поведение было отмечено и в естественных условиях. Джейн Гудолл, долгое время наблюдавшая за поведением шимпанзе в природе, описывает случай, когда в группе шимпанзе, за которой она наблюдала, появилась самка‑каннибалка, Пэшн, поедавшая чужих детенышей. Самке Мифф удалось спасти своего детеныша от Пэшн, и впоследствии, когда она встретилась с Пэшн не один на один, а в компании дружественных самцов, Мифф выказала сильное возбуждение и смогла донести до самцов идею, что Пэшн ей очень не нравится и ее надо наказать – по крайней мере, самцы, увидев поведение Мифф, устроили Пэшн агрессивную демонстрацию73 .
Можно предполагать, что во всех таких случаях обезьяны передают не столько сам конкретный опыт, сколько свои эмоции по его поводу. И, вероятно, в большинстве случаев этого бывает достаточно, поскольку антропоиды способны очень тонко различать нюансы того, что психологи называют “невербальной коммуникацией”. Например, шимпанзе Уошо смогла угадать, что работавшие с ней Роджер и Дебора Футс – муж и жена, хотя они намеренно старались на работе вести себя друг с другом не как супруги, а как коллеги. “Никто не сравнится с шимпанзе в умении понимать невербальные сигналы!” – написал по этому поводу Р. Футс74 .
Однако, если информация, которую необходимо передать, достаточно необычна, такой способ коммуникации дает сбои. Так, в описанном выше примере объяснить, что в точности произошло, Мифф не смогла – в противном случае самцы бы, наверное, не ограничились демонстрацией, а выгнали бы Пэшн из группы или, по крайней мере, предупредили бы об опасности дружественных им самок.
Впрочем, когда в языковых проектах обезьяны получают в свое распоряжение более совершенное коммуникативное средство – язык‑посредник (и, к слову, более понятливого собеседника – человека), они оказываются в состоянии облечь свой собственный опыт и взгляды на мир в знаковую форму (см. примеры в гл. 1).
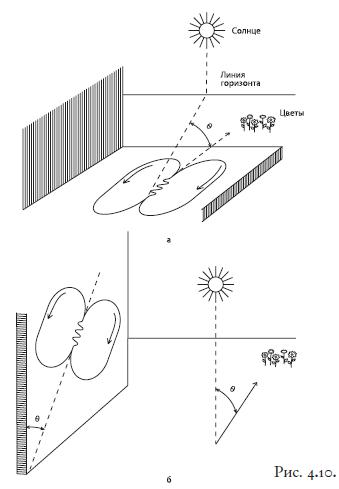
Рис. 4.10. Виляющий танец.
Попытки расшифровать коммуникативные системы животных предпринимались неоднократно. Одна из наиболее успешных – расшифровка виляющего танца медоносной пчелы австрийским биологом Карлом фон Фришем75 . Угол между осью танца и вертикалью (если пчела танцует на вертикальной стенке) соответствует углу между направлением на пищу и направлением на Солнце, продолжительность движения пчелы по прямой несет информацию о расстоянии до источника корма; кроме того, имеют значение скорость, с которой движется пчела, виляние брюшком, движение из стороны в сторону, звуковая составляющая танца и т. д. – всего по меньшей мере одиннадцать параметров. Блестящим подтверждением правильности этой расшифровки стала созданная Акселем Михельсеном76 пчела‑робот: ее танцы в улье (см. фото 17 на вклейке), управляемые компьютерной программой, успешно мобилизовывали пчел‑сборщиц на поиски корма. Пчелы правильно определяли направление на кормушку и расстояние до нее – даже несмотря на то, что пчела‑робот не давала сборщицам запаховой информации.
Но многие другие коммуникативные системы оказались сложнее. Так, не удалось в точности выяснить, какие движения муравьев, прикасающихся антеннами к своим сородичам, информируют их, скажем, о повороте направо. У дельфинов удалось определить лишь “свист‑подпись”. Единственный расшифрованный сигнал волков – “звук одиночества”. Гудолл77 отмечает, что шимпанзе издают звук “хуу” “только при виде небольшой змеи, неизвестного шевелящегося создания или мертвого животного”, – но почти ни про какие другие звуки шимпанзе ничего столь же определенного пока сказать нельзя.
Широко известны опыты Эмила Мензела78 с шимпанзе. Экспериментатор показывал одному из шимпанзе тайник со спрятанными фруктами, и потом, когда обезьяна возвращалась к своей группе, она неким образом “сообщала” соплеменникам о местонахождении тайника – по крайней мере, те отправлялись на поиски, явно имея представление о том, в каком направлении следует идти, и даже иногда обгоняли сообщавшего. Если одному шимпанзе показывали тайник с фруктами, а другому – с овощами, группа не колеблясь выбирала первый тайник. Если в тайнике была спрятана игрушечная змея, шимпанзе приближались к нему с некоторой опаской. Но как именно шимпанзе передавали соответствующую информацию, так и осталось загадкой. Высокоранговые особи, казалось, не делали для этого вовсе ничего, но тем не менее добивались понимания, низкоранговые, напротив, разыгрывали целую пантомиму, делали выразительные жесты в соответствующем направлении – но все равно мобилизовать группу на поиски тайника им не удавалось.
Для расшифровки смысла того или иного сигнала необходимо, чтобы его появление взаимно‑однозначно соответствовало либо некоторой ситуации во внешнем мире, либо строго определенной реакции особей, воспринимающих сигнал. Поэтому так легко оказалось расшифровать систему предупреждения об опасности у верветок: крик с определенными акустическими характеристиками (отличными от характеристик других криков) жестко коррелирует (а) с наличием леопарда в зоне видимости и (б) с убеганием всех слышащих сигнал обезьян на тонкие верхние ветки.
Но большинство сигналов волков, дельфинов, шимпанзе таких жестких корреляций не обнаруживают. Как отмечает Е.Н. Панов, они могут “в разное время выступать в разных качествах”79 . Например, у шимпанзе один и тот же сигнал оказывается связан и с ситуацией дружелюбия, и с ситуацией подчинения, и даже с ситуацией агрессии. По мнению Панова, это свидетельствует о том, что с точки зрения теории информации “эти сигналы существенно вырождены”80 и никакого внятного смысла не имеют. Но то же самое рассуждение применимо и ко многим выражениям человеческого языка. Если рассматривать слова не в словаре, где каждому из них приписана вполне определенная семантика, а в составе выражений, произносимых в реальных жизненных ситуациях, легко видеть, что они, подобно сигналам животных, могут в разное время выступать в разных качествах. Например, предложение “Молодец!” может выступать и в качестве похвалы (“Уже все уроки сделал? Молодец!”), и в качестве порицания (“Разбил чашку? Моло‑дец!”). Слово “точка” может обозначать начало (“точка отсчета”) и конец (“на этом поставим точку”), маленький черный кружок, изображенный на бумаге (“проведите прямую через точку А и точку В”), и реальное, подчас довольно большое и не всегда круглое место (“торговая точка”). Таким образом, если следовать логике Е.Н. Панова, человеческий язык тоже, пожалуй, придется признать вырожденным с точки зрения теории информации.
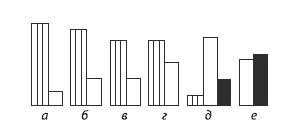
Рис. 4.11. Эти шесть сигналов шимпанзе (выделенных этологом Яаном ван Хооффом) могут, хотя и с различной частотой, выступать в разных ситуациях – и при дружелюбном взаимодействии (заштрихованные столбцы), и для демонстрации подчинения (белые столбцы), и при агрессии (черные столбцы). Относительная высота столбцов отражает частоту, с которой каждый сигнал был зафиксирован в соответствующей ситуации. Сигнал “визг с оскаленными зубами” (д) используется во всех трех типах взаимодействий 81 .
В человеческих языках не существует, видимо, ни одного выражения, которое бы вызывало всякий раз одну и ту же реакцию. Даже услышав крик “Пожар!”, одни люди бросятся участвовать в спасении, другие – мародерствовать, третьи станут созерцать происходящее, не предпринимая никаких действий, а четвертые просто пройдут мимо. Как писал Тютчев, “Нам не дано предугадать…”. Не существует и ситуации, которая бы однозначно вызывала появление того или иного сигнала, – люди по‑разному строят свои высказывания в зависимости от того, какие элементы ситуации представляются им в данном конкретном случае более важными, учитывают тот фонд знаний, которым, по их представлениям, обладает слушающий, отражают в высказывании свое отношение к ситуации (а нередко и к слушающему) и т. д., и т. д.{32}. Колоссальная избыточность, которой обладает любой чел
Дата добавления: 2016-02-02; просмотров: 50887;