Был ли язык у австралопитека? 2 страница
Генетики же до самого недавнего времени были уверены, что метисации не было и не могло быть, поскольку неандертальские последовательности нуклеотидов отсутствуют в митохондриальной ДНК населения Европы – того региона, где обитали классические неандертальцы54 . Впрочем, в статье, написанной генетиком А. Темплтоном еще в 2005 г.55 , утверждается, что данные митохондриальной ДНК не могут являться однозначным свидетельством; разные участки разных молекул ДНК хранят следы разных событий в истории человечества: и следы заселения Евразии архантропами около 1,9 млн. лет назад, и следы более поздней ашельской экспансии (650 тыс. лет назад), и следы миграции из Африки человека разумного около 100 тыс. лет назад. Вероятность того, что смешения между различными представителями рода Homo не было, составляет, по его подсчетам, 10–17. В пользу метисации свидетельствуют и данные приматологов: многие виды обезьян “живут в тесном контакте друг с другом, обитают в сходных биотопах и зачастую образуют смешанные группы или стада”56 , в таких группах появляются межвидовые гибриды, практически все из которых плодовиты57 (засвидетельствованы гибриды павианов анубисов и павианов гамадрилов, многих видов макак; гибриды бонобо с шимпанзе и гориллами58 ). Любопытно, что существование в составе смешанных стад может способствовать “значительному расширению коммуникативного репертуара”, по крайней мере, у одного из видов59 .
С прочтением ядерного генома неандертальца стало окончательно ясно: неандертальцы и сапиенсы скрещивались между собой (хотя и не слишком активно), у жителей современной Евразии имеется 1–4 % неандертальских генов60 .
Коммуникативная система не оставляет непосредственных следов, которые могли бы быть обнаружены в ходе раскопок, поэтому о наличии языка у тех или иных ископаемых видов (точнее, о возможных свойствах их коммуникативной системы) приходится судить по косвенным признакам. Чаще всего в качестве определяющего свидетельства существования языка называют опущенную гортань: она представляет собой ту высокую цену, которую наши предки заплатили за возможность овладения членораздельной звучащей речью. Но гортань не содержит костей, а мягкие ткани не сохраняются, поэтому все имеющиеся в науке сведения о положении гортани у того или иного вида гоминид – это реконструкции, выполненные на основании предположения, что положение гортани связано с величиной так называемого базикраниального угла (угла основания черепа): чем сильнее изогнуто основание черепа, тем ниже расположена гортань61 . У австралопитеков и Homo habilis изгиб основания черепа примерно такой же, как у современных человекообразных обезьян, у архантропов он выражен значительно сильнее – поэтому некоторые исследователи (например, А.А. Зубов) предполагают, что “первыми “говорящими” гоминидами были, возможно, Homo ergaster ”62 . Поскольку у неандертальца перегиб основания черепа был меньше, чем у гейдельбергского человека, можно предположить, что “неандертальская специализация” была сопряжена с уменьшением приспособленности к речи. Впрочем, недавние исследования указывают на отсутствие (у современных людей) надежной корреляции между величиной изгиба базикраниума и положением гортани63 .
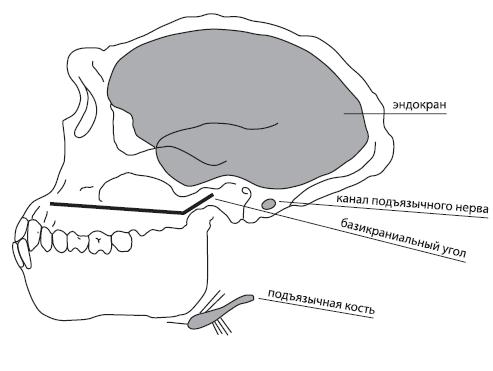
Рис. 3.5. Анатомические характеристики (предположительно имеющие отношение к языку), которые можно наблюдать на ископаемых останках 64 .
Кроме того, следует уточнить, что для членораздельной речи важно не положение гортани само по себе, а соотношение длины ротовой полости и длины глотки: у современного человека эти длины приблизительно одинаковы, так что части языка, расположенные во рту и в глотке, примерно уравниваются, и это дает возможность четко различать в произношении все звуки, включая “крайние” гласные – [i], [u] и [a]65 . У других видов гоминид такой возможности, видимо, не было, поскольку они, как видно по сохранившимся черепам, обладали достаточно сильно выступавшими вперед челюстями и, соответственно, длинной ротовой полостью. Чтобы уравновесить такую длину рта соответствующей длиной глотки, гортань должна была бы располагаться где‑то в груди, что крайне маловероятно66 . Впрочем, невозможность произнесения “крайних” гласных сама по себе не может свидетельствовать ни об отсутствии языка (язык мог в принципе быть и жестовым), ни даже об отсутствии членораздельной звучащей речи – просто, если таковая была, она должна была в большей степени полагаться на различия согласных звуков67 .
Помимо гортани, ниже, чем у других приматов, у человека расположена подъязычная кость – и это тоже связано с расширением возможностей для звучащей речи (см. гл. 2). Как была расположена эта кость у других представителей клады человека – неизвестно, поскольку эта небольшая кость, не прикрепленная к остальному скелету, обычно не сохраняется. К настоящему времени исследователи располагают лишь очень немногими образцами подъязычной кости. Наиболее известны найденная в Израиле (пещера Кебара) подъязычная кость неандертальца68 и подъязычная кость гейдельбергского человека из Испании (район Атапуэрка, пещера Сима де лос Уэсос); кроме того, в пещере Сидрон в Астурии, близ Пилона, была найдена частично сохранившаяся подъязычная кость гоминида, принадлежавшего к неандертальцам (или гейдельбержцам)69 . Все эти кости, хотя и несколько отличаются по своему строению, чрезвычайно сходны с теми, которые можно наблюдать у современных людей70 (в частности, в них отсутствуют отверстия для воздушных мешков, свойственных современным шимпанзе71 ), и это дает возможность высказать гипотезу, что голосовые аппараты неандертальца, гейдельбержца и современного человека анатомически чрезвычайно близки. Напротив, найденная недавно подъязычная кость афарского австралопитека оказалась такой же, как у шимпанзе72 . Однако вопрос о том, может ли форма подъязычной кости служить надежным индикатором наличия речи у соответствующего вида, остается дискуссионным73 .
Подбородочный выступ имеется лишь у неоантропа, но у некоторых неандертальцев прослеживается определенная тенденция к его формированию74 . Присутствует в зачаточном состоянии подбородочный выступ, например, у неандертальца из Кебары (экземпляр Кебара 2, известный как “Моше”), обладателя сохранившейся подъязычной кости. Правда, из этого едва ли можно делать какие‑либо выводы о системе коммуникации. Возможно, более показательно строение внутренней поверхности нижней челюсти75 : в ее середине (в области симфиза) у человека имеются подбородочные ости (места прикрепления подбородочно‑язычной мышцы); у обезьян же на этом месте ямка (поскольку эта мышца прикрепляется к кости не сухожилием, а мясистой частью). Челюсти ископаемых гоминид демонстрируют спектр переходных форм.
Предположения о наличии у разных видов гоминид звучащей речи делаются также на основании толщины канала подъязычного нерва. Как заметили Ричард Кей, Мэт Картмилл и Мишель Балоу76 , у представителей рода Homo этот канал как абсолютно, так и относительно больше, чем у человекообразных обезьян и австралопитековых. Однако, как уже говорилось в главе 2, и эта характеристика не является достаточно показательной.
Возможно, более информативна ширина позвоночного канала: она позволяет судить о возможности тонкой регулировки дыхания, очень важной для речи, по крайней мере, если эта речь не ограничивается односложными высказываниями. По имеющимся данным, у неандертальца и гейдельбергского человека этот канал был примерно таким же широким, как у неоантропа, тогда как у архантропа и даже у Homo antecessor – значительно уже77 . Увеличение ширины позвоночного канала не является простым следствием прямохождения: об этом говорит тот факт, что вполне прямоходящие архантропы имели позвоночный канал примерно такой же ширины, как у приматов, не входящих в кладу человека78 .
Косвенным свидетельством адаптации к звучащей речи является устройство органов слуха. Как показали Игнасио Мартинес и его коллеги, уже у гейдельбергского человека слуховой анализатор был настроен на преимущественное распознавание звуков на тех частотах, которые характерны для речи современного человека. Поскольку эти гоминиды являются не прямыми предками нашего вида, а представителями линии, ведущей к неандертальцам, можно предполагать, что эти же частоты использовал для своей звуковой коммуникации наш общий с неандертальцами предок79 .
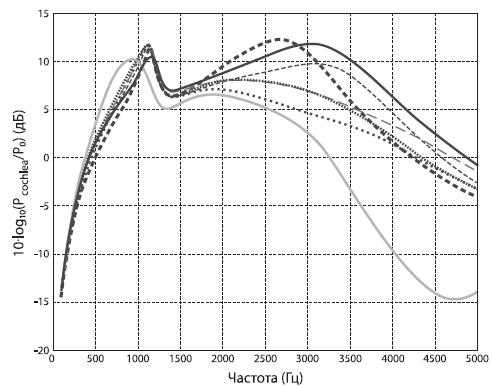
Рис. 3.6. Пунктирные линии – слух различных экземпляров Homo heidelbergensis (из местонахождения Сима де лос Уэсос); для сравнения: сплошная черная линия – слух сапиенса, серая – шимпанзе 81 .
Целый ряд гипотез о происхождении языка отводит важную роль в этом процессе развитию прямохождения80. Например, по мнению Э. Карстейрса‑Маккарти, человеческий язык вполне может оказаться “лишь побочным продуктом изменения речевого аппарата, вызванного переходом наших предков к двуногому образу жизни”82.
За последнее время картина анатомического строения предков человека пополнилась новыми деталями, больше стало известно и о походке различных Homo и австралопитековых; обсуждаются различные гипотезы о причинах перехода к двуногой локомоции83 . Существует несколько концепций того, какие адаптивные преимущества давала бипе‑дия. Уильям Леонард в своем обзоре84 пишет: “Оуэн Лавджой (C. Owen Lovejoy) из Кентского университета предположил, что передвижение на двух ногах освободило австралопитекам руки для ношения детей и собирания корма. Позднее Кевин Хант (Kevin D. Hunt) из Индианского университета высказал мнение, что бипедия возникла как особая поза во время кормежки, благодаря которой гоминидам стала доступна пища, прежде находившаяся вне пределов их досягаемости. А Питер Уилер (Peter Wheeler) из Университета Джона Мора в Ливерпуле заявил, что прямохождение (благодаря которому сократилась площадь поверхности тела, доступная воздействию обжигающего африканского солнца) позволило нашим предкам лучше регулировать температуру тела. Проведенные мной исследования наводят на мысль, что двуногая походка оказалась энергетически более выгодной, чем четвероногая. Главные факторы, влияющие на энергозатраты при движении, – масса тела и скорость передвижения. Как ни удивительно, но при скорости, сравнимой со скоростью ходьбы, двуногая походка гораздо экономичнее, чем квадрупедия” (т. е. передвижение на четырех конечностях). Среди представителей клады человека, которые были всеядными существами и вели “широкий пищевой поиск”85 , отбор по признаку энергетической эффективности передвижения должен был играть существенную роль.
Существует, впрочем, и такая точка зрения, согласно которой бипедия представляла собой вынужденное свойство: приматы, хорошо адаптированные к вертикальному древолазанию и/или брахиации (это такой способ передвижения по ветвям, при котором особь висит на верхних конечностях и перемещается, попеременно цепляясь ими то за одну ветку, то за другую), просто не смогли двигаться по земле с опорой на четыре конечности, хотя прямохождение приносило на первых порах больше вреда, чем пользы86 .
Ходить на двух ногах гоминиды умели, по‑видимому, уже очень давно – это надежно подтверждают подробные исследования останков Ardipithecus ramidus 87 . Достаточно хорошо сохранившийся скелет самки ардипитека (получившей прозвище “Арди”) демонстрирует сочетание “обезьяньих” (ориентированных на эффективное лазание по деревьям) и “человеческих” (ориентированных на прямохождение) признаков. Впрочем, не исключено, что бипедия возникла еще раньше – судя по положению затылочного отверстия, на двух ногах передвигался сахелантроп; некоторые приспособления к бипедии отмечены у оррорина и у Ardipithecus kadabba 88 .
Было также установлено, что Арди не опиралась при ходьбе на костяшки пальцев, как это делают современные шимпанзе и гориллы – кисти ее рук совершенно не приспособлены для этого. Зато они гораздо лучше подходят для тонких манипуляций с предметами (а следовательно, для орудийной деятельности), чем руки современных человекообразных обезьян.
После исследования Арди стало ясно, что сценарий, приведенный на шуточном рисунке в начале главы, в корне неверен: ходьба с опорой на костяшки пальцев – это не тот способ передвижения, которым пользовались предки человека, а специализация боковых ветвей – шимпанзе и горилл, – их способ уйти от прямохождения.
Для афарского австралопитека, как пишет У. Леонард, “были характерны такие признаки двуногого существа, как сводчатая стопа, непротивопоставляющийся большой палец стопы и некоторые особенности строения колена и таза”89 (тем самым, его походка была более уверенной, чем у ардипитека). Прямоходящими были африканские архантропы90 (недавно этот вывод был подтвержден находкой ископаемых следов в Кении, на берегу озера Туркана91 ), а также гейдельбержцы и неандертальцы92 .

Рис. 3.7. Один из следов архантропа, обнаруженных в Кении (в 5 км от деревни Илерет) 93 . Справа, для сравнения, след современного человека.
Приспособления к двуногому передвижению не обнаруживают корреляции с теми анатомическими характеристиками, которые считаются важными для звучащей речи, – величиной базикраниального угла, толщиной позвоночного канала и т. д. Кроме того, ходить на двух ногах – причем так же, как люди, полностью разгибая колени, – умеют, как недавно выяснилось, бесспорно не имеющие языка орангутаны94 (см. фото 7 на вклейке). Так что, скорее всего, прямого влияния на становление языка бипедия не оказала. Впрочем, освобождение передних конечностей для манипулятивной активности дало возможность сильно расширить спектр поведенческих стратегий, повысив тем самым “спрос” на развитые средства коммуникации. Кроме того, “вертикальное положение позвоночника дает возможность обладания бóльшим объемом мозга, чем горизонтальное или наклонное при равных размерах тела”95 , а больший мозг дает возможность иметь более совершенную коммуникативную систему. Хождение на двух ногах привело также к расширению общего поля зрения, облегчило возможность обращать внимание сородичей на те или иные факторы окружающей среды – а это очень важно для языка, который, по свидетельству психологов, используется прежде всего не для побуждений, а именно для обращения внимания (англ. sharing attention )96 . Это обусловило возможность возникновения в дальнейшем так называемого “остенсивного определения” – определения при помощи указательного жеста, который имеется во всех человеческих культурах97 (хотя в целом ряде их считается невежливым) {24}.

Рис. 3.8. Это остенсивное определение.
Для обеспечения функционирования языка необходим развитый мозг, и в кладе человека наблюдается постепенное увеличение его объема. Ранние гоминиды по этому показателю были сравнимы с шимпанзе: мозг сахелантропа имел объем98 320–380 см3, ардипитека – 300–350 см3, австралопитека афарского – 380–450 см3, мозг Australopithecus africanus – 440–450 см3 (по некоторым данным, около 500 см3), мозг Paranthropus boisei – 530 см3 (ср. 350–550 см3 у шимпанзе). Но уже у Homo habilis мозг был объемом 500–800 см3(в среднем 650 см3), примерно таким же был мозг Homo rudolfensis (775 см3) и Homo georgicus (600–680 см3). Существенно увеличился мозг у архантропов – 900–1100 см3, еще более – у гейдельбержцев (например, индивид, найденный на стоянке Вертешсёллёш, имел мозг объемом 1300 см3, индивид из пещеры Петралона – 1170 см3, индивид из пещеры Фонтешевад, известный как Фонтешевад II, – 1450 см3). У неандертальцев объем мозга достигал примерно 1400–1600 см3, что даже несколько превышает среднюю величину, характерную для современного человека, – 1350–1400 см3. Впрочем, у многих ископаемых неоантропов мозг был крупнее. Например, у кроманьонцев из грота Кро‑Маньон объем мозга определяется как 1590 см3 (Кро‑Маньон I), 1450 см3 (Кро‑Маньон II) и 1775 см3, (Кро‑Маньон III), у индивида Схул V – 1518 см3, у Омо 1 – 1400 см3. Для сравнения: неандерталец Ля Ферраси 1 имел мозг объемом 1681 см3, неандерталец Шанидар 1 – 1610 см3, неандерталец Амуд I – 1750 см3.
Следует, однако, отметить, что даже для хорошо сохранившихся черепов оценки объема мозга у разных исследователей сильно варьируют. Так, объем мозга неандертальца из Ля Шапелль‑о‑Сен оценивается в диапазоне от 1093 до 1626 кубических сантиметров100 – и это при том, что измерения проводились по практически не требующему реконструкции черепу. А, например, от экземпляра Неандерталь 1 осталась лишь черепная крышка – соответственно, подсчеты объема его мозга оказываются еще более гипотетическими {25}.
Наиболее точные размеры мозга можно получить путем измерения эндокрана – отлива внутренней полости черепа (см. прежде всего работы Вероники Ивановны Кочетковой, Ральфа Холлоуэя и Станислава Владимировича Дробышевского101 ). При этом следует иметь в виду, что, как отмечает С.В. Дробышевский, “эндокран не полностью соответствует форме мозга, поскольку между последним и черепом человека располагаются мозговые оболочки”102 . Подсчеты, проведенные на основании измерений черепа, менее точны. Но, каковы бы ни были конкретные цифры, можно с уверенностью утверждать, что общая тенденция к увеличению размеров мозга в ходе эволюции гоминид несомненно имела место.
Как отмечает Т. Дикон103 , поскольку метаболизм одного нейрона ограничивает число синапсов, которые он может поддерживать, то, чем больше мозг, тем меньший процент от общего числа его нейронов будет связан с каждым данным нейроном. Таким образом, рост мозга ведет к фрагментации его функционирования и потере скорости обработки информации (поскольку разные структуры, необходимые для ее обработки, находятся физически дальше друг от друга и связываются через бóльшее количество промежуточных узлов). Зато такой мозг позволяет увеличить объем хранимой и различаемой информации. С ростом мозга растет способность применять знания к новым контекстам, но прямой зависимости между размером мозга и умом нет104 . Общеизвестно, что мозг И.С. Тургенева был сильно больше средней величины, характерной для современного человека (2012 г.), а мозг Анатоля Франса – сильно меньше (1017 г.), но это не мешало им обоим быть великими писателями.
Существенно также, что наш мозг способен расти после рождения, увеличиваясь ко взрослому состоянию более чем втрое; судя по недавним исследованиям, особенности и темпы роста мозга у Homo sapiens отличаются от того, что имело место у других гоминид, в частности, у неандертальцев105 .
С энергетической точки зрения мозг – не самый выгодный орган. Будучи равным всего 2 % веса тела человека, он потребляет около 20–25 % всей энергии организма (у взрослого человека в состоянии покоя, а у новорожденного – даже 60 %), что значительно больше, чем у обезьян (около 8 %)106 . Значит, если у того или иного вида гоминид наблюдается рост мозга, можно с уверенностью утверждать, что эти гоминиды производили существенно больше энергии, чем их предшественники, и могли часть этой энергии тратить на поддержание функционирования коммуникативной системы. Расчеты У. Леонарда показывают, что “типичный австралопитек с массой тела 40–43 кг и объемом мозга 450 куб. см расходовал на его нужды примерно 11 % энергии. А Н. erectus , весивший 63–66 кг и обладавший мозгом в 850 куб. см, ежедневно затрачивал 250 из 1500 ккал, т. е. 16 % энергии”107 . С другой стороны, более развитый мозг может обрабатывать больше информации, а значит, повышается спрос на источники этой информации, в том числе на коммуникативные действия сородичей.
Английским антропологом Артуром Кизсом108 в начале ХХ века была выдвинута гипотеза “мозгового рубикона”, согласно которой достижение “подлинно человеческого” культурного уровня и даже, возможно, появление речи у тех или иных гоминид определяется тем, достиг ли объем мозга цифры 750 см3 (кстати, именно таков в среднем объем мозга у детей того возраста, в котором происходит овладение речью). Если бы это было так, первыми обладателями языка были бы архантропы. Но открытие “хоббитов” поставило на этой гипотезе крест: типичные архантропы по своим анатомическим характеристикам, эти гоминиды умели делать орудия, пользоваться огнем, охотиться на карликовых слонов – стегодонов109 . Но их мозг был уменьшен пропорционально уменьшению тела и составлял примерно 400 см3 (это в границах объема, характерного для шимпанзе, однако структура мозга у “хоббитов” была совершенно иной, существенно более прогрессивной110 ). Если такой культурный уровень соответствует наличию речи, значит, Homo floresiensis умели говорить, – но тогда это должно означать, что для существования языка достаточно куда меньшего объема мозга, чем предполагает гипотеза “мозгового рубикона”. Если же предположить, что “хоббиты” говорить не умели, это будет означать, что “подлинно человеческий” культурный уровень достижим и без языка – и тогда гипотеза “мозгового рубикона” лишается оснований. Впрочем, аргументированная критика этой гипотезы высказывалась и ранее111 .
Для языка имеет значение не только абсолютный объем мозга, но и его структура. На эндокране можно увидеть, какие зоны мозга существовали у различных видов предков человека, установить их форму, а также узнать, какие зоны мозга увеличены у исследуемого вида по сравнению с его предком. Так, например, по эндокрану австралопитека афарского прослеживается увеличение задней теменной ассоциативной коры. Это “указывает на наличие естественного отбора на более сложные формы познавательного поведения, чем те, которые наблюдаются у шимпанзе”112 . Как пишет С.В. Дробышевский, “на основании того, что мы знаем о функциях разных долей мозга современного человека, можно предположить, что грацильные австралопитеки, вероятно, обладали несколько большей чувствительностью и подвижностью рук, чем современные человекообразные обезьяны. В сравнении же с массивными австралопитеками грацильные, вероятно, имели более развитый зрительный анализатор”113 . Согласно данным Ф. Тобайаса114 , у Homo habilis по сравнению с австралопитековыми значительно расширяется теменная область, появляется подобный человеческому узор борозд, а также возникают хорошо выраженные специфические выпуклости “в областях, где у людей находятся речевые центры (поле Брока и поле Вернике)”115 . По наблюдениям С.В. Дробышевского, у “ранних Homo ” (т. е. Homo habilis и Homo rudolfensis ) строение мозга было чрезвычайно изменчиво. Но при этом общим для всех изученных эндокранов “является прогрессивное развитие области, которая у современного человека отвечает за согласование устной речи и движений рук”116 .
У архантропов, помимо формирования рельефа в области зоны Брока, происходит “бурное развитие зрительных центров затылочной доли”117 , что, по мнению Дробышевского, связано с началом выхода в открытые саванны. Кроме того, у них, “судя по развитию теменной доли, значительно развились тактильные возможности и намного улучшилась координация движений”118 – и это позволяло им производить орудия более совершенного, ашельского, типа. Из архаичных особенностей строения мозга стоит отметить то, что “лобная доля мозга была низкая и узкая”119 .
У гейдельбергского человека “росла высота лобной доли, а расширение этой доли, вероятно, несколько обгоняло расширение всего мозга”120 , при этом “височная доля… имела уже вполне современные пропорции, хотя и меньшие, чем у современного человека, размеры”121 . В целом “форма мозга Homo heidelbergensis свидетельствует о резком прогрессе в области контроля за движениями, в том числе способностей к прогнозированию и планированию своих будущих действий. Необходимо также отметить бурное развитие области, обеспечивающей согласование речи и движений рук, а также рельефное выступание зоны Брока”122 .
У неандертальца “ширина мозга крайне велика, максимальна для всех групп гоминид. Очень характерны относительно малые размеры лобной и теменной долей при очень большой – затылочной. В орбитной области (на месте зоны Брока) были развиты рельефные бугры”123 . В частности, значительное развитие зоны Брока отмечается у “грацильных” неандертальцев типа Ортю. По мнению С.В. Дробышевско‑го, мозг неандертальца “отличался от мозга современного человека, вероятно, бóльшим развитием подкорковых центров подсознательного контроля за эмоциями и памятью, но, вместе с тем, меньшим сознательным контролем за этими же функциями”124 . Вероятно, они также несколько отставали от современного человека “в плане возможностей контроля и координации движений”125 .
У неоантропа сильно увеличиваются зоны синтеза сигналов, абстрактного мышления126 . Возрастание способностей к сложной психической и интеллектуальной деятельности демонстрирует рост лобных долей (в особенности префронтальных формаций коры). Как отмечает Дробышевский, “у людей верхнего палеолита, в сравнении с неандертальцами, увеличилась длина и высота мозга, выросла лобная доля”127 . И это позволило представителям нашего вида делать выводы и выбирать поведенческие программы с учетом большего числа факторов. Соответственно, у неоантропа возникла потребность в такой коммуникативной системе, которая сможет предоставить эти факторы в его распоряжение. Гоминиды с меньшими лобными долями получили бы меньшую выгоду от высокоразвитого языка.
Нередко одним из наиболее надежных свидетельств наличия языка у тех или иных ископаемых гоминид считается наличие в их мозге зоны Брока (а также зоны Вернике)128 . Однако следует отметить, что эндокран показывает лишь форму того или иного участка мозга, но не его функцию; функция же может меняться не только в ходе эволюции, но и в течение жизни индивида, поскольку нейроны способны перепрофилироваться. Гомологи человеческих речевых центров, как уже говорилось выше (гл. 2), имеются и у обезьян, так что вопрос о том, был ли язык у того или иного ископаемого вида гоминид, анализ эндокранов полностью не проясняет.
Сильно возрос за последнее время объем археологических данных о культурах и технологиях эпохи палеолита, – а по мнению некоторых исследователей, например, археолога Иайна Дэвидсона, это едва ли не единственное, что может пролить свет на происхождение языка.
Нередко считается, что сами по себе орудия являются свидетельством “обратного моделирования”: ментальный образ объекта предшествует его появлению в реальном мире129 . Как пишет А.А. Зубов, “созданная мозгом абстрактная модель предмета или процесса воплощается в жизнь, то есть как бы встраивается в реальный мир, где ее до этого не было и, часто, не могло быть”130 . Согласно чеканной формулировке английского археолога Гордона Чайлда, “ручное рубило как стандартизованное орудие есть само по себе ископаемая концепция”. Такой подход восходит к гипотезе, высказанной еще в XVIII в. эдинбургским профессором Джеймсом Барнетом Монбоддо, согласно которой использование орудий является надежным критерием, выделяющим человека из мира животных.
Орудийная деятельность связывалась также с развитием доминации правой руки, которая делает орудия, – именно поэтому, как считается, основным для языка является, как и для правой руки, левое полушарие. Однако, как показывают исследования (см. гл. 2), связь языка с доминированием правой руки, вопреки распространенному мнению, не прямая, а опосредованная131 .
Дата добавления: 2016-02-02; просмотров: 1218;