Был ли язык у австралопитека? 1 страница
При исследовании происхождения языка невозможно обойти вниманием вопрос о том, у кого и когда появился язык. Возник ли он вместе с видом Человек разумный или, может быть, лишь на каком‑то этапе его существования? Или, может быть, он появился уже у архантропов, а какими‑то зачатками языка обладали еще австралопитеки?

Рис. 3.1. Эволюция человека (шуточный вариант).
Знания об ископаемых гоминидах за последнее время значительно пополнились 1 . Только за первые годы ΧΧΙ в. были открыты такие новые виды, как Ardipithecus kadabba 2 , Kemanthropus platyops 3 , Orrorin tugenensis 4 , Sahelanthropus tchadensis 5 , Homo floresiensis 6 ; совсем недавно был описан вид Australopithecus sediba 7 ; кроме того, была пересмотрена классификация уже известных видов. С этим связан довольно значительный разнобой в назывании одних и тех же гоминид в разных работах, например, один и тот же вид может быть назван и зинджантроп, и Australopithecus boisei , и Paranthropus boisei ; под термином Homo erectus может пониматься любой из питекантропов / синантропов, но ряд исследователей выделяет наиболее ранних эректусов в отдельный вид Homo ergaster (или подвид Homo erectus ergaster ). Человека современного физического типа одни считают отдельным видом Homo sapiens , другие – подвидом Homo sapiens sapiens , наряду с неандертальцем (который в этом случае обозначается как Homo sapiens neanderthalensis ).
Все виды, которые после того, как 7–8 млн. лет назад линия, ведущая к человеку, отделилась от линии, ведущей к современным человекообразным обезьянам, оказались на первой из этих линий, составляют так называемую “кладу человека”. Всех представителей этой клады я буду, следуя работе Л.Б. Вишняцкого8 , обозначать словом “гоминиды”, пользоваться термином “австралопитековые” для всех австралопитеков и парантропов (а также ардипитеков, кениантропа, са‑хелантропа и оррорина), термином “архантропы” для Homo ergaster и Homo erectus и термином “неоантропы” для людей современного физического типа.
Стало уже окончательно ясно, что эволюция человека не может быть описана как простая линейная последовательность – разные виды сосуществовали и даже, возможно, конкурировали друг с другом. Например, современниками человека разумного, появившегося более 100 тыс. лет назад, были неандертальцы9 (последние находки датируются временем 28–29 тыс. лет назад), а также Homo floresiensis (жившие 95–12 тыс. лет назад)10 . Парантропы жили в то же время, что и Homo rudolfensis, Homo habilis , а возможно, и Homo erectus . Было даже высказано мнение, что “наша состоявшаяся эволюционная история – это только один из многих потенциально существовавших ее сценариев, который, в отличие от других, не остался запасным лишь в силу во многом случайного стечения мало связанных между собой обстоятельств”11 .
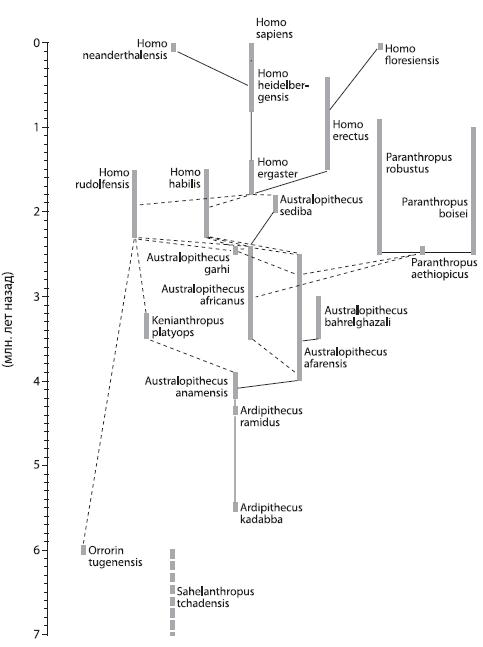
Рис. 3.2. Эволюционное древо человека (одна из существующих версий). Линии обозначают связи между предками и потомками (спорные случаи отмечены пунктиром).
У истоков генеалогического древа человека стоят гоминиды конца миоцена. Наиболее древние находки относятся к видам Sahelanthropus tchadensis {23} (7–6 млн. лет назад; датировки приведены по данным сайта Антропогенез. ру12 ), Orrorin tugenensis (6 млн. лет назад) и Ardipithecus kadabba (5,5 млн. лет назад); по мнению эфиопского палеоантрополога Йоханнеса Хайле‑Селассие и его коллег, все эти три вида достаточно близки друг к другу и, возможно, не должны считаться отдельными родами13 .



От Ardipithecus kadabba происходит Ardipithecus ramidus 14 (4,4 млн. лет назад), считающийся предком всех австралопитеков и парантропов: от него происходит самый примитивный из австралопитеков – Australopithecus anamensis (4,2–3,9 млн. лет назад), предок афарского австралопитека (Australopithecus afarensis – вид, к которому относится найденная Дональдом Джохансоном знаменитая “Люси”; время существования – 4,0–2,5 млн. лет назад).

Потомки афарского австралопитека – Australopithecu safricanus (3,5–2,4 млн. лет назад), Australopithecus bahrelghazali (3,5–3,0 млн. лет назад) и Australopithecus garhi (2,5 млн. лет назад). От австралопитека афарского (или, по другим версиям, от австралопитека африканского) ведут свой род парантропы (они же “массивные австралопитеки” – в противоположность прочим, “грацильным”): Paranthropus aethiopicus (2,5 млн. лет назад) и его потомки Paranthropus robustus (2,5–0,9 млн. лет назад) и Paranthropus boisei (2,5–1,0 млн. лет назад). От какого‑то из грацильных австралопитеков – по одной версии, от Australopithecus afarensis , по другой, от A. africanus , возможно, через промежуточную ступень A. garhi – происходит первый человек, Homo habilis (“человек умелый”; впрочем, некоторые ученые считают его не первым человеком, а, скорее, последним австралопитеком15 ), предок архантропов. В ряде работ часть хабилисов выделяют в отдельный вид – Homo rudolfensis (2,3–1,5 млн. лет назад), причем этот последний иногда рассматривается как переходное звено к Homo habilis в узком смысле (2,3–1,5 млн. лет назад) от австралопитека афарского16 . Французский антрополог Брижит Сеню, нашедшая вместе со своими коллегами оррорина, считает, что Homo rudolfensis должен быть объединен с Orrorin tugenensis в род Praeanthropus , который ведет прямо к человеку, минуя австралопитеков.


А по мнению Мив Лики, нашедшей и описавшей в 1999 г. вид Kenianthropus platyops (3,5–3,2 млн. лет назад), Homo rudolfensis – прямой потомок кениантропа и должен быть включен в род Kenianthropus , ведущий свое происхождение от Australopithecus anamensis . По мнению южноафриканского палеоантрополога Филиппа Тобайаса, Homo rudolfensis , как и Homo habilis , является потомком Australopithecus africanus ; существует и точка зрения, возводящая Homo rudolfensis вместе с Homohabilis к Australopithecus afarensis . Было высказано также предположение о происхождении Homo habilis (или Homo rudolfensis ) от Australopithecus sediba (2,0–1,8 млн. лет назад), потомка Australopithecus africanus 17 . Как пишет палеонтолог Александр Владимирович Марков, “распутать этот клубок трудно, потому что все формы, о которых идет речь, – и поздние австралопитеки, и ранние люди – приходились друг другу очень близкой родней (примерно как современные шимпанзе и бонобо). По‑видимому, в период между 2,5 и 1,8 млн. лет назад в Южной и Восточной Африке жили разнообразные популяции продвинутых гоминид, подвергавшиеся сходному действию отбора и эволюционировавшие более или менее в одном направлении, но с разной скоростью. Какие‑то из этих популяций скрещивались друг с другом, какие‑то нет. Подразделить эту разношерстную компанию на “еще обезьян” (австралопитеков) и “уже людей” (ранних Homo ) можно лишь с большой долей условности”18 .



От Homo habilis (или, по другим версиям, от Homo rudolfensis или даже напрямую от Australopithecus sediba 19 ) происходят архантропы, Homo erectus (1,5 млн. лет назад – 400 тыс. лет назад, а возможно, если верна датировка яванских находок и эти находки не относятся к виду Homo heidelbergensis , даже 50 тыс. лет назад). Переходная форма была обнаружена в пещере Сварткранс (Южная Африка), это экземпляр, известный под названием SK 80/846/847 (или “Телантроп III ”20 ). Возможно, правильнее выделять его в отдельный вид – Homo ergaster ; останки сходного типа были найдены и в других местах; вид датируется временем 1,8–1,4 млн. лет назад. Еще одна возможная переходная форма между хабилисами и архантропами – гоминиды, жившие в районе нынешнего Дманиси (Грузия) 1,8 млн. лет назад. Обычно их относят к Homo erectus или Homo ergaster , но иногда рассматривают и как отдельный вид – Homo georgicus 21 . Высказывалось предположение, что эволюция африканских и азиатских архантропов проходила независимо (немецкий антрополог Франц Вейденрейх на основании особенностей строения зубов предположил, что азиатские популяции неоантропов произошли непосредственно от азиатских Homo erectus , см. ниже). Отдельная ветвь архантропов – “хоббиты” с острова Флорес (Индонезия), Homo floresiensis (95–12 тыс. лет назад), имевшие рост около метра и объем мозга около 400 см3. Скептики, правда, считают, что найденные останки принадлежат представителю нашего вида, больному микроцефалией (при этой врожденной патологии резко уменьшается объем черепа). Но сторонники реальности вида Homo floresiensis указывают на то, что, помимо найденного в 2003 г. единственного целого черепа (известного как Лянг Буа, или LB1), в 2004 г. была обнаружена также челюсть столь же маленького размера, принадлежавшая, судя по степени истертости зубов, взрослой особи22 . В пользу реальности “хоббита” говорят и данные посткраниального скелета (так называются все кости, кроме черепа): как показали исследования, по строению плечевой кости “хоббит” гораздо больше похож не на современного человека, а на кенийского Homo erectus (экземпляр, известный как “подросток из Нариокотоме”, “Turkana boy” или KNM‑WT 15000), его стопа отличается по строению от стопы современного человека24 , а запястье более примитивно не только чем у неоантропа или неандертальца, но даже чем у их общего предка25 . Недавние исследования слепка мозговой полости черепа Лянг Буа 1 показали, что мозг “хоббитов” был устроен не так, как у больных микроцефалией26 .
Первооткрыватели “хоббитов” Майкл Морвуд, Питер Браун и их коллеги считают, что эти гоминиды, вероятнее всего, происходят от Homo erectus , попавших на остров около 800 тыс. лет назад, – такую дату дают первые каменные

Рис. 3.3. Homo erectus
орудия, найденные на Флоресе (если это действительно орудия, а не природные образования27 ), весьма похожие на те, что были найдены вместе с “хоббитами”28 . Впрочем, кости Homo erectus , которые должны были бы изготавливать эти орудия, на острове Флорес не найдены. Высказывалось предположение, что предками “хоббитов” могли быть не эректу‑сы, а более примитивные гоминиды, подобные тем, что были найдены в Дманиси29 .

Следующая ступень эволюции человека – Homo heidelbergensis (800–200 тыс. лет назад). Антрополог Александр Александрович Зубов, подробно описавший этот вид, отмечает у него мозаику черт, свойственных архантропам и неоантропам (но не неандертальцам). Объем мозговой коробки гейдельбержца был сравнительно невелик, меньше, чем у неандертальца, но высота свода при этом была
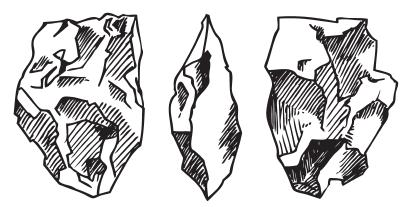
Рис. 3.4. Вот такими камнями, возможно, пользовались предки “хоббитов”, жившие на о. Флорес около 800 тыс. лет назад 30 .
больше и лоб был менее покатым. Затылок был не вытянутым назад, как у неандертальца (“неандертальский шиньон”), а более округлым, как у сапиенса. Более изогнутым было основание черепа – если связывать это с положением гортани (см. ниже), то получается, что к членораздельной звучащей речи гейдельбержцы были ближе, чем неандертальцы. В то же время у гейдельбержцев отмечаются и архаичные признаки – кости свода черепа очень толстые, толще, чем у неоантропа, а заглазничная область сильно сужена31 . Многие образцы, относимые ныне к этому виду, ранее рассматривались либо как прогрессивные эректусы, либо как ранние неандертальцы. Переходным звеном от архантропов к гейдельбержцу является, вероятно, группа гоминид, представленная экземпляром OH9 (иногда называемым “шелльским человеком”, или Homo leakey ), который был найден в слое II в Олдувайском ущелье32 и датируется временем 0,9–1 млн. лет назад33 .
Близки к Homo heidelbergensis останки, обнаруженные в пещере Гран Долина (район Атапуэрка) в Пиренеях и датируемые временем порядка 800 тыс. лет назад. Испанские исследователи Хуан Луис Арсуага, Антонио Росас и Хосе Бермудес‑де‑Кастро (авторы находки) полагают34 , что это отдельный вид (названный ими Homo antecessor ), являющийся непосредственным предком человека разумного. Гейдельбергский человек, по их мнению, также является потомком, Homo antecessor .

А.А. Зубов рассматривает Homo antecessor как “представителя примитивной формы Homo heidelbergensis или еще более раннего, неопределенного вида”35 .
Гейдельбержцы широко расселились по земному шару: их находят в Европе, Африке (как Северной, так и Южной) и Азии – в Израиле, Карабахе, Индии, Китае (эти находки некоторые исследователи считают результатом эволюции местных, китайских, архантропов) и даже на Яве.
А.А. Зубов считает гейдельбергского человека предком и неандертальца, и человека разумного, но при этом оговаривается, что вся европейская линия вела к неандертальцу, неоантропы же произошли от популяций, оставшихся в Африке (эти последние иногда выделяют в отдельный вид Homo helmei 36 , а иногда причисляют к виду Homo antecessor ).
Далее, в процессе адаптации к условиям приледниковой зоны развиваются анатомические особенности, характерные для неандертальца (см. картину Николая Ковалева на вклейке, илл. 10): небольшой рост (примерно 160 см у мужчин), плотное телосложение, энергетически выгодный теплообмен, высокий уровень физического развития. Мощная мускулатура кисти руки обеспечивала огромную силу захвата и удара. Наиболее ранние неандертальцы – так называемые “атипичные”, вероятно, представляют собой переходную ступень от Homo heidlbergensis к неандертальцу37 . “Классические” неандертальцы, обладавшие полным комплексом неандерталоидных черт, датируются временем около 70–30 тысяч лет назад.
Долгое время считалось, что неандертальцы были предками неоантропов – эта точка зрения в рамках стадиальной концепции обосновывалась тем, что по степени сложности орудий неандертальцы занимают промежуточное положение между архантропами и сапиенсами времен верхнепалеолитической революции. Но впоследствии оказалось, что неандертальцы и неоантропы в течение многих десятков тысяч лет были современниками, а значит, первые не могли быть предками вторых. Последнюю точку в этом вопросе поставили генетики. В 1997 г. Сванте Пяябо и его коллеги38 сумели прочитать фрагмент митохондриальной ДНК неандертальца (найденного еще в середине XIX в. близ Дюссельдорфа). Митохондриальная ДНК – сравнительно небольшая кольцевая молекула дезоксирибонуклеиновой кислоты, которая находится не в ядре клетки, а в митохондриях, которые передаются человеку только от матери. Некоторые участки мтДНК (так же, как и некоторые участки ядерной ДНК) не кодируют, согласно современным данным, ничего такого, что могло бы подпасть под действие естественного отбора, поэтому замены нуклеотидов на этих участках происходят чисто случайно, и, соответственно, по скорости этих замен можно вычислить время, когда жила последняя общая “праматерь” рассматриваемых организмов. С тех пор были проанализированы митохондриальные ДНК еще нескольких неандертальцев39 (в том числе неандертальского ребенка, чьи останки были найдены на Кавказе, в пещере Мезмайская40 ), а в 2008 г. был полностью прочитан митохондриальный геном неандертальца из Хорватии, жившего 38 тыс. лет назад (экземпляр Виндия 75)41 . Генетический материал неандертальцев сравнили с материалом современных людей и шимпанзе. Выяснилось, что все рассмотренные неандертальцы достаточно близки друг к другу – примерно так же, как некоторые популяции современных людей, и гораздо ближе, чем разные популяции шимпанзе. Расстояние от шимпанзе до неандертальцев оказалось примерно таким же, как расстояние от шимпанзе до современных людей, различие же между неандертальцами и современными сапиенсами указывает на то, что последний их общий предок жил 660 000 ± 140 000 лет назад. Значит, неандерталец не был предком неоантропа и, соответственно, неандертальская система коммуникации – какова бы она ни была – не является системой, непосредственно предшествующей человеческому языку. К моменту встречи с неандертальцами сапиенсы уже, несомненно, обладали некоторой коммуникативной системой, сформировавшейся на базе коммуникативной системы вида‑предка; вопрос же о том, оказали ли на нее какое‑либо влияние контакты с неандертальцами, остается открытым. Впрочем, на мой взгляд, такое влияние едва ли имело место, поскольку языки Европы (т. е. той территории, где могли происходить наиболее интенсивные контакты с неандертальцами) отличаются от всех остальных языков мира не в большей степени, чем отличаются от других языки любого другого крупного региона. В случае же неандертальского влияния ожидалось бы, что языки Европы будут противопоставлены прочим языкам сильнее, чем любые из этих прочих языков противопоставлены друг другу.
Что же касается современных людей, то их общая прародительница (названная “митохондриальной Евой”) была определена еще раньше. По мтДНК, полученной от 147 представителей различных рас (африканцев, европейцев, азиатов, австралийцев и жителей Новой Гвинеи), американский генетик Алан Уилсон из Калифорнийского университета и его коллеги установили, что “митохондриальная Ева” жила более 100 тыс. лет назад. В настоящее время в литературе фигурируют разные датировки общей прародительницы человечества по генетическим данным – от приблизительно двухсот до 130 тыс. лет назад. Примерно такие же датировки были получены на основании анализа Y‑хромосомы, которая, наоборот, передается только по мужской линии: в качестве наиболее вероятного времени существования “Y‑хромосомного Адама” называют период от 57 до 135 (а по некоторым данным – до 180) тыс. лет назад. Разумеется, это не означает, что все люди произошли от одной пары – для нормального воспроизводства необходима полноценная популяция (минимально необходимое количество особей для разных видов различно), просто если через какое‑то время никто из дочерей (внучек, пра‑пра‑…‑правнучек) прародительницы не оставит потомков женского пола, ее мтДНК будет утрачена; точно так же утратится Y‑хромосома, если ни у кого из мужчин‑потомков (не важно, в каком поколении) мужчины‑предка не будет сыновей. Несколько групп генетиков независимо пришли к выводу, что людей в популяции “митохондриальной Евы” было в те времена от 10 до 30 тысяч, то есть Homo на этом этапе своего существования прошли через так называемое “бутылочное горлышко” (сильное сокращение численности).
Генетика помогла также прояснить вопрос о том, было ли происхождение человека нашего вида единичным процессом или же неоантропы возникли в нескольких местах независимо. В середине прошлого века и гипотеза моноцентризма, и гипотеза полицентризма имели своих сторонников. Основным аргументом в пользу гипотезы полицентризма служило сходство в строении зубов синантропов и современных жителей Юго‑Восточной Азии, отмеченное Ф. Вейденрейхом42 . Их так называемые “лопатообразные резцы” не являются приспособлением к условиям окружающей среды, а значит, едва ли могли возникнуть независимо. Существовала также гипотеза “широкого моноцентризма”, выдвинутая советским ученым Яковом Яковлевичем Рогинским – согласно ей, центр сапиентации был один, но охватывал очень большую территорию.
Но, по современным данным, обе эти гипотезы неверны. Анализ мтДНК позволяет определить степень родства разных групп людей между собой и построить генеалогическое древо современного человечества. Первое ветвление на этом древе указывает на Африку – одна из групп африканцев выделилась раньше, чем прочие группы (включающие население всего остального мира, в том числе и других африканцев) разделились между собой. Это говорит о том, что человек разумный возник всего один раз – в Африке, и, судя по малочисленности предковой популяции, территория, на которой это происходило, была достаточно невелика.
На Африку указывают и ископаемые данные. Как пишет А.А. Зубов, именно там “спектр переходных форм в процессе сапиентации не имеет четких разрывов в последовательности”43 . Самый отдаленный предок современных людей, более всего близкий к архантропам, был найден в Бодо, его существование датируется временем 600 тыс. лет назад. Следующую ступень представляет собой индивид, чьи останки известны под названием Омо 2, далее – Элие Спрингс, потом – индивид Летоли 18, и наконец, Омо 1, чьи анатомические характеристики при самой строгой оценке не выходят за рамки внутривидового разнообразия человека разумного. Все эти находки были сделаны в Африке. Они демонстрируют мозаику архаичных и прогрессивных черт, так что одни и те же экземпляры разные исследователи причисляют к разным видам. Индивид из Бодо в одних работах фигурирует как “ранний архаичный сапиенс”, в других – как гейдельбергский человек44 ; Омо 2 называют и просто сапиенсом, и поздним архаичным сапиенсом, и гейдельбержцем, а в некоторых работах он и все последующие члены этого ряда, кроме Омо 1, называются отдельным видом Homo helmei .
Несколько нарушают гармонию этого построения датировки: согласно современным данным, индивид Элие Спрингс жил 100–200 тыс. лет назад, Летоли 18 – 120 ± 30 тыс. лет назад. Для Омо 1, первого бесспорного представителя нашего вида, в литературе нередко фигурирует датировка 130 тыс. лет назад, полученная по раковинам нильских устриц, найденных чуть выше него. Но недавно45 в Эфиопии на месте его находки были проведены дополнительные исследования. Омо 1 (как и несколько более архаичный Омо 2) был найден между двумя слоями вулканического пепла. Нижний из этих слоев расположен прямо под костеносным слоем, его возраст, определенный калий‑аргоновым методом, составляет 196 ± 2 тыс. лет. Верхний слой пепла залегает значительно выше костей и датируется временем 104 ± 1 тыс. лет назад. По совокупности данных оба черепа получили датировку 195 ± 5 тыс. лет назад. Таким образом, если все приведенные датировки верны и классификационная принадлежность всех останков определена правильно, придется признать, что приведенный выше ряд переходных форм от архантропа к неоантропу представляет собой не лестницу последовательно переходящих друг в друга ступеней‑стадий, а, как и во многих других случаях эволюции, спектр одновременно существовавших вариантов, из которых в конце концов остался только один.
Итак, уже около 200 тыс. лет назад в Африке жили анатомически современные люди, и гипотезы о происхождении языка непременно должны это учитывать.
В частности, эти датировки показывают, что при исследовании происхождения языка не приходится, увы, полагаться на достижения сравнительно‑исторического языкознания. Наиболее хорошо изученный праязык – индоевропейский – отстоит от нас лишь на 6 тысячелетий. Примерно таким же временем – в рамках первого десятка тысяч лет до наших дней – датируется и время распада таких известных праязыков, как уральский, алтайский, картвельский, северокавказский, сино‑тибетский. Следующий уровень реконструкции – макросемьи, в частности, ностратическая (куда включаются индоевропейские, уральские, алтайские, дравидийские, картвельские и, предположительно, ряд других языков) и сино‑кавказская (объединяющая северокавказские языки с сино‑тибетскими, енисейскими и, возможно, еще некоторыми). Распад этих праязыков датируется временем порядка 14 тысяч лет назад. Весьма вероятно, что некоторые из существующих макросемей родственны между собой – но даже столь смелая гипотеза продвигает нас в глубь времен не более, чем на 20 тысячелетий46 . Представители американской школы компаративистики (школы Джозефа Гринберга) утверждают, что смогли классифицировать все существующие на Земле языки и свести их к одному общему предку47 , но уровень доказательности этой гипотезы совершенно недостаточен48 . Таким образом, попытки экстраполировать на время глоттогенеза данные, полученные в рамках сравнительно‑исторического языкознания, при нынешнем уровне знаний едва ли могут претендовать на достоверность.
Но даже если праязык человечества будет когда‑нибудь надежно реконструирован, это, скорее всего, не приблизит нас к пониманию глоттогенеза: реконструкции доступны лишь те языки, которые не имеют принципиальных отличий от ныне существующих, так что в реконструированном всеобщем праязыке будут обнаружены слова, состоящие из фонем, – а не, скажем, “диффузные выкрики”. Происхождение же этих слов из элементов предшествующей языку коммуникативной системы останется вне рамок сравнительно‑исторического исследования.
Один из “вечных”, не решенных пока вопросов сравнительно‑исторического языкознания – так называемая “проблема моногенеза‑полигенеза”: произошли ли все известные языки Земли от одного общего праязыка или от нескольких разных. Но к возникновению языка из не‑языка эта проблема прямого отношения не имеет. В том случае, если язык возник в нескольких местах независимо (а это могло быть как в случае наличия нескольких независимых очагов сапиентации, так и в случае, если язык возник уже после появления – безразлично, в одном месте или в нескольких – человека разумного), один из этих праязыков человечества мог успеть вытеснить все остальные, оставшись на какой‑то момент единственным, и дать начало всем известным (ныне существующим и письменно зафиксированным вымершим) языкам. В том же случае, если очаг возникновения языка был один, остается возможность, что те черты, которые приводятся (например, в работах лингвиста А.Н. Барулина49 ) как аргументы в пользу полигенеза, были заимствованы в языки человека разумного из до‑языковых (но неизбежно близких к языку) коммуникативных систем – все эти черты относятся к числу таких, которые могут проникнуть в язык при интенсивных контактах50 .
Другой, едва ли не более важный, вопрос состоит в том, могли ли другие представители рода Homo внести свой вклад в генофонд вида Homo sapiens , – если да, то это повышало бы вероятность того, что человеческий язык хранит следы влияния коммуникативных систем других гоминид. О возможности скрещивания между разными представителями рода Homo (прежде всего между неоантропами и неандертальцами) в литературе идут жаркие дискуссии. Археологи и антропологи усматривают у многих останков как неандерталоидные, так и сапиентные признаки – таковы неандерталоиды Палестины, поздние (фазы IVB и V периода Вюрм II, т. е. порядка 30 тыс. лет назад) “грацильные” (или “микродонтные”, по терминологии Мари‑Антуанетт де Люмле51 ) неандертальцы Ортю (Франция) и Каригуэлы (Испания). По мнению одних исследователей, они представляют собой переходный этап от неандертальца к неоантропу, другие делают на основании анализа этих останков вывод о возможности метисации (или гибридизации, как предпочитают говорить те, кто считает их разными видами) между неандертальцами и сапиенсами52 ; третьи не исключают, что перед нами что‑то вроде попытки природы создать на основе неандертальца альтернативный сапиенсу разумный вид53 .
Дата добавления: 2016-02-02; просмотров: 992;