Механізм передачі збудження в хімічних синапсах
Ідея гуморальної передачі нервового імпульсу є порівняно давньою. Перша згадка про хімічну медіацію належить Дю Буа Раймону (1877). У 1921 році віденським фармакологом Отто Льові проведено знаменитий дослід, яким остаточно було стверджено хімічну передачу збудження в синапсі (рис. 2.3).
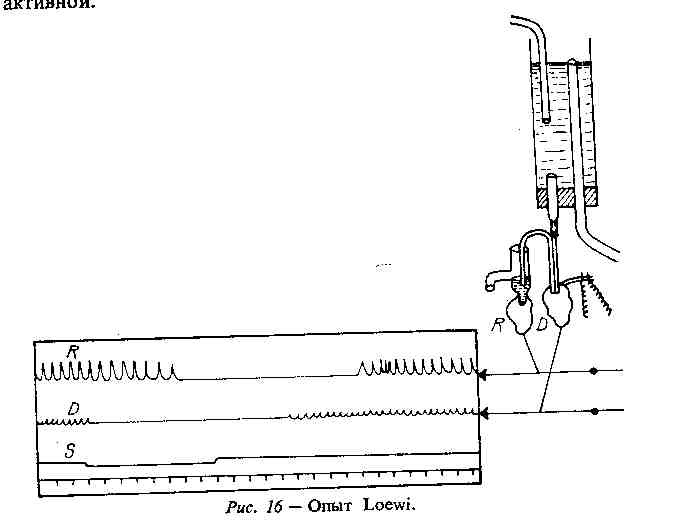 Рис. 2.3. Дослід О.Льові (Loewwi) у модифікації Кана.
Рис. 2.3. Дослід О.Льові (Loewwi) у модифікації Кана.
О.Льовi подразнював блукаючий нерв iзольованого серця жаби. Серце уповiльнювало свою роботу. Потiм омиваючий розчин переносився з цього серця до iншого, також iзольованого, яке не стимулювалось: друге серце давало таку ж реакцiю. Було зроблено висновок, що при подразненнi блу- каючого нерва першого серця в поживну рiдину переходить певний медiатор.
Тривалий «вагусний» ефект на ізольоване серце було отримано після додавання в перфузат рос- линного алкалоїду – езерину, який є блокатором холінестерази. Атропіном блокувалась дія блукаю- чого нерва на ізольоване серце. Так було доведе- но, що хімічним передатчиком збудження з блукаю-
чого нерва на серце є ацетилхолін (Льові, Навратіл, 1926). Пізніше, за пропозицією чеського дослідника Р.Кана, було зконструйовано дворогу канюлю, до якої приєднувались два ізольованих серця: одне з вом, блукаючим або симпатичним, а друге – без нього. Отже, поживний розчин (перфузат) був загальним для обох сердець. У такому разі при подразненні нервів (блукаючого чи симпатичного) першого серця, у другому серці відбувались аналогічні зміни діяльності – відповідно уповільнення або прискорення скоро- чень.
Хiмiчний механiзм передачi збудження в верхньому шийному симпатичному ганглiї продемонстрував О.В.Кiбяков (1933). Використавши мiкроелектродну технiку для внутрiшньоклiтинної реєстрацiї синаптич- них потенцiалiв нейронiв ЦНС, Дж. Екклс зробив висновок про хiмiчну природу передачi збудження в си- напсах спинного мозку.
Стосовно хімічної медіації симпатичних волокон, знов таки Льові (1921), користуючись описаною спериментальною моделлю, навів перший прямий доказ звільнення гуморального фактора, який володіє серцевоприскорюючою дією, і назвав його “Accelerans-stoff”. Завдяки використанню флюорометричних методів кількісного виявлення катехоламінів (адреналіну і норадреналіну) у тканинах і рідинах організму, фон Ейлер (1946-1947) довів, що очищені витяжки симпатичних нервів і робочих органів містять майже виключно норадреналін. Таким чином було зроблено уточнення, що головним хімічним медіатором постгангліонарних симпатичних закінчень є норадреналін, а адреналін відіграє скоріше роль гормону наднирників. Тепер встановлено, що норадреналін, як медіатор, присутній в корі мозку, гіпоталамусі, сто- вбурі мозку, мозочку, спинному мозку.
 Серед медіаторів є різні хiмiчнi ни. До них належать: ацетилхолiн, ламiни (адреналiн, норадреналiн, дофамiн), серотонiн, нейтральнi амiнокислоти (глутамiнова, аспарагiнова), кислi амiнокислоти (глiцин, γ-амiномасляна кис- лота – ГАМК), полiпептиди (речовина Р, енкефалiн, соматостатин та iн.), iншi речовини (АТФ, гiстамiн, простагланди- ни). Вiдповiдно синапси класифiкують за типом медiатора як холiнергiчнi, адре- нергiчнi та iн.
Серед медіаторів є різні хiмiчнi ни. До них належать: ацетилхолiн, ламiни (адреналiн, норадреналiн, дофамiн), серотонiн, нейтральнi амiнокислоти (глутамiнова, аспарагiнова), кислi амiнокислоти (глiцин, γ-амiномасляна кис- лота – ГАМК), полiпептиди (речовина Р, енкефалiн, соматостатин та iн.), iншi речовини (АТФ, гiстамiн, простагланди- ни). Вiдповiдно синапси класифiкують за типом медiатора як холiнергiчнi, адре- нергiчнi та iн.
Медiатор синтезується в тiлi нейрона. Звiдси вiн транспортується по аксону до синаптичних закiнчень – кiнцевих бляшок, де i накопичується у везикулах. Видiлення може вiдбуватись як спонтанно, без зовнiшньої стимуляцiї у станi вiдносного спокою, так i при збудженнi (рис. 2.4).
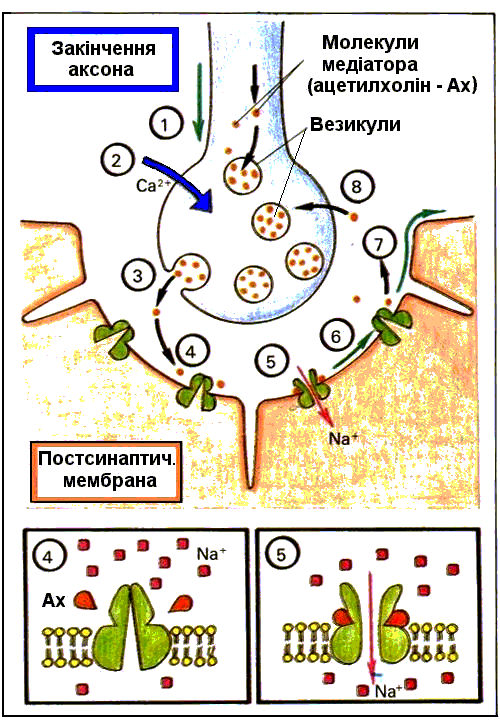
Рис. 2.4. Механізм передачі збудження в синапсі.
1. Надходження потенціалу дії до пресинаптичної частини синапсу. 2. Вхід iонiв кальцiю у кiнцеву бляшку. 3. Видiлення у синап- тичну щiлину кванта медіатора (ацетилхоліна) і його дифузія через внутрiшньощiлинну речовину до постсинаптичної частини. 4. Ацетилхолiн дiє на особливо чутливi до нього дiлянки – рецеп- тивну субстанцію каналу. 5. Постсинаптична мембрана на короткий час стає проникною для iонiв, насамперед для натрiю (дещо й для кальцію) і у постсинаптичнiй мембранi виникає деполяризацiя. 6. Виникнення на постсинап- тичнiй мембранi деполяризацiйного потенцiалу – збуджувальний постсинаптичний потенцiал (ЗПСП). 7. Руйнування ацетилхолiну холiнестеразою; рецептори повертаються у
вихiдний стан. 8. Всмоктування продуктів розщеплення медіатора в пресинаптичну мембрану.
А.Фетт i Б.Катц (1952) встановили, що коли в пресинаптичному вiддiлi руйнується одна везикула, то звiльняється вiд 6 до 10 тис. молекул ацетилхоліну. Цю кiлькiсть було названо квантом медiатора. При подразненнi нерва в пресинаптичнiй частинi синапсу одночасно руйнується вiд 250 до 500 везикул, у синаптичну щiлину видiляється відповідна кiлькiсть квантiв медіатора. Процес звiльнення медiатора запускається ПД, що надходить до аксо- на, за участю iонiв кальцiю, котрi входять через пресинаптичну мембрану i сприяють виходу медіатора в синаптичну щiлину. Далi медіатор дифундує через внутрiшньощiлинну речовину до постсинаптичної час- тини, де дiє на особливо чутливi до нього дiлянки – рецептивну субстанцію.
Унаслiдок дiї ацетилхолiну на холiнорецептори постсинаптична мембрана на короткий час стає про- никною для iонiв, насамперед для натрiю (дещо й для кальцію). У постсинаптичнiй мембранi виникає деполяризацiя. Одного кванта медiатора досить для зменшення потенцiалу на 0,5 мВ. Такий потенцiал зветься мiнiатюрним потенцiалом. При одночасному звiльненнi 250-500 квантiв ацетилхолiну (2,5-5 млн. молекул) спостерiгається максимальне збiльшення кiлькостi мiнiатюрних потенцiалiв. Мiнiатюрнi потенцiали здатнi до сумації, унаслiдок чого на постсинаптичнiй мембранi утворюється деполяризацiйний потенцiал. Вiн пов'язаний з поступовим пiдвищенням проникностi натрiю через постсинаптичну мембрану з щiлини всередину другого нейрона (його сому або волокно) i має всi властивостi локального потенцiалу. Його назва – збуджувальний постсинаптичний потенцiал (ЗПСП). Коли проникнiсть натрiю (у зв'язку зi збiльшенням кiлькостi медiатора) зростає, ЗПСП досягає максимальної амплiтуди – критичного рiвня деполяризацiї (КРД), що складає 15 мВ. Це і є передумовою виникнення ПД (тобто збудження), який розповсюджується по всiй поверхнi нейрона. Для виникнення ПД необхiдно, щоб ЗПСП виник не менш нiж у 50 синапсах. У цьому випадку ЗПСП досягне критичного рiвня. Тривалість ЗПСП в периферичних і більшості нервових синапсів майже однакова: фаза зростання – 2 мс, спаду – 10-15 мс. Найбiльша
збудливiсть у мембрани початкового сегмента аксона (аксонного горбика) завдяки найвищій щільності Na+-каналів на 1 мм2 клітинної мембрани та його оголеності – відсутності мієлінової оболонки, отже й зменшеному опору. Саме тут започатковується ПД, який потiм розповсюджується по аксону й охоплює
тiло клiтини.
У синаптичнiй щiлинi медіатор (наприклад, ацетилхолiн) знаходиться дуже короткий промiжок часу (1-2 мс). Тут вiн руйнується відповідним ферментом (холiнестеразою). Рецептори повертаються у вихiдний стан, а продукти розщеплення медіатора в значнiй кiлькостi всмоктуються пре- i постсинаптичними мем- бранами і ресинтезуються у везикулах.
Пiсля завершення ПД у багатьох нейронах ЦНС спостерiгається вiдносно довга слiдова гiперполяризацiя. Це пояснюється тим, що мембрана клiтини на вiдмiну вiд аксонiв має значну кiлькiсть кальцiєвих каналiв. Пiд час деполяризацiї цi канали активуються. Iони кальцiю, якi входять в середину клiтини, активують зворотну проникнiсть мембрани для калiю, що й спричиняє слiдову гiперполяризацiю. Цей механiзм вiдiграє важливу роль в регуляцiї частоти ПД нервової клiтини.
У фармакологічній практиці відомий ряд речовин, котрі запобігають виникненню збудження в синапсах. Це можна здійснити різними шляхами:
1) заблокувати проведення збудження до пресинаптичної мембрани (місцеві анестетики);
2) заблокувати виділення медіатора (зменшити концентрацію Са2+, ввести Мn2+, Mg2+, які блокують проникливість Са2+ через пресинаптичну мембрану, отже й ініціацію виділення медіатора);
3) порушити синтез ацетилхоліну в синапсах (геміхоліній);
4) заблокувати холінорецептори постсинаптичної мембрани (атропін; зміїна отрута (бунгаротоксин); кураре - конку- рент ацетилхоліну, який зв’язує холінорецептори в нервово-м’язових синапсах; декаметоній, сукцинілхолін - виклика- ють деполяризацію пост-синаптичної мембрани і таким чином інактивують Na+);
5) загальмувати дію холінестерази (медіатор діє довго й викликає стійку деполяризацію).
Згідно принципу Дейла, кожен нейрон здатний виділяти лише один якийсь медіатор. Останнім часом знайдено відхилення від такої закономірності: визначено нейрони, кожен з яких спроможний виробляти декілька медіаторів.
 Крім іонотропних медіаторів існують також метаболотропні. Здебільшого це нейропептиди. Свій вплив на постси- наптичну мембрану вони здійснюють шляхом активації специфічних ферментів мембрани. Як наслідок, у нейронах активуються вторинні посередники (месендже-
Крім іонотропних медіаторів існують також метаболотропні. Здебільшого це нейропептиди. Свій вплив на постси- наптичну мембрану вони здійснюють шляхом активації специфічних ферментів мембрани. Як наслідок, у нейронах активуються вторинні посередники (месендже-
ри), які в свою чергу запускають каскади внутрішньоклітинних процесів, тим самим впли- ваючи на функцію клітин.
До месенджерів відносять такі 4 системи: 1) аденілатциклаза – цАМФ; 2) гуанілатциклаза – цГМФ; 3) фосфоліпаза С – інозітол – трифосфат;
4) іонізований кальцій. Вони забезпечують вплив як на іонну проникливість мембран, так і на син- тез та виділення медіаторів; регулюють також синтез білків, енергетичний обмін.
Дата добавления: 2015-12-11; просмотров: 3423;