Синапси гальмiвної дiї
Гальмування– активний процес в ЦНС, який проявляється пригнiченням збудження або повним його припиненням у вiдповiдь на подразнення.
Довгий час iснувала думка, що для нервової системi можливий тiльки процес збудження, а пригнiчення фiзiологiчної реакцiї пов'язане зi зменшенням процесу збудження. Вiдкриття про- цесу гальмування як фiзiологiчного феномена було зроблене при дослiдженнi впливу блукаю- чого нерва на скорочення серця (брати Вебер, 1845). Явище центрального гальмування вiдкрив I.М.Сєченов у 1863 р. Він довiв, що подразнення структур стовбура мозку (низхідний відділ рети- кулярної формації) викликає пригнiчення спин- номозкових центрiв жаби (рис. 2.5).
Рис. 2.5. Схема центрального гальму- вання за І.М.Сєченовим
На схемі показано розповсюдження нервових імпульсів від гальмівних нейронів стовбура мозку до спинного мозку при накладанні кристалика NaCl на ділянку зорових горбів.
Гальмування за рахунок центрів спинного мозку встановив Гольц. Відтоді починається вивчення галь-
мування як самостiйного нервового процесу, який викликається збудженням i проявляється пригнiченням iншого збудження.
На вiдмiну вiд процесу збудження, який виявляється в двох головних формах – ПД, здатного до роз- повсюдження та локальних потенцiалiв, гальмування може виникати тiльки у виглядi локального процесу й завжди пов'язане з iснуванням специфiчних гальмiвних синапсiв. Пресинаптичнi закiнчення гальмiвних синапсiв належать аксонам гальмiвних нейронiв, якi пригнiчують активнiсть усiх нервових клiтин, з котри- ми вступають у синаптичний контакт. Прикладом гальмiвних нейронiв у спинному мозку є нейрони Рен- шоу, у головному мозку – нейрони Пуркiн'є кори мозочка.
Сучаснi електрофiзiологiчнi дослiдження дозволили встановити два принципово рiзнi способи галь- мування клiтин: постсинаптичне гальмування (зниження збудливостi соми чи дендритiв нейрона) та пресинаптичне гальмування (зменшення чи припинення видiлення медiатора пресинаптичним нервовим закiнченням) (рис. 2.6).
 Рис. 2.6. Гальмування в ЦНС.
Рис. 2.6. Гальмування в ЦНС.
Гальмівні нейрони заштриховано.
А – пресинаптичне гальмування: пригнiчується видiлення медiатора в збуджувальних синапсах. Структурною основою такого процесу гальмування є аксо-аксональнi синапси, якi утворюються аксонами гальмiвних вставних нейронiв i аксо- нальними закiнченнями збуджувальних нейронiв. Активацiя аксо-аксонального синапса супроводжується деполяризацiєю пресинаптичної мембрани за типом катодичної депресiї, тобто натрiєвої iнактивацiї, до чого причетна кальцiєва проникнiсть. При надходженнi до такої деполяризованої дiлянки ПД затримується або зменшується його амплітуда у порівнянні з нормальною i тому у відповідному синапсi не видiляється медiатор.
Б – постсинаптичне гальмування:ПД закiнчення аксона, що належить до гальмiвного нейрона, викликає видiлення медiатора, який активує калiєвi канали, і як наслідок виникає гiперполяризацiя постсинаптичної мембрани – гальмiвний пост- синаптичний потенцiал (ГПСП).
Постсинаптичне гальмування розвивається насту- пним чином: ПД закiнчення аксона, що належить до гальмiвного нейрона, викликає видiлення медiатора, який
активує калiєвi канали. Виникає гiперполяризацiя постсинаптичної мембрани – гальмiвний постсинапти-
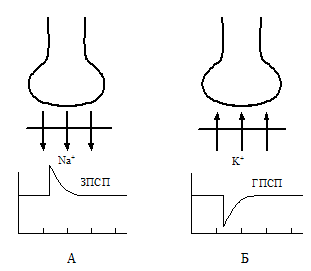 чний потенцiал (ГПСП) (рис. 2.7 ).
чний потенцiал (ГПСП) (рис. 2.7 ).
Рис. 2.7. Механізми генерації збуджувального пос- тсинаптичного потенціалу (ЗПСП - А) та гальмівно- го постсинаптичного потенціалу (ГПСП - Б).
При розвитку ГПСП підвищується критичний рівень деполяризації (КРД), у зв’язку з чим пригнiчується дiяльнiсть натрiєвих каналiв i зменшується можливiсть розвитку процесу деполяризацiї, тобто ЗПСП. Такий механізм гальмування властивий здебільшого аксо- дендритним синапсам. У розвитковi ГПСП аксо- соматичних синапсів провідну роль вiдiграють iони хлору, якi, проникаючи всередину нейрона, також при- зводять до гiперполяризації мембрани. У спинному мозку гальмiвним медiатором може бути глiцин. Його блокатором (конкурентом на субсинаптичній мембрані) є стрихнін.
Пресинаптичне гальмування полягає в тому, що пригнiчується видiлення медiатора в збуджувальних
синапсах. Структурною основою такого процесу гальмування є аксо-аксональнi синапси, якi утворюються аксонами гальмiвних вставних нейронiв i аксональними закiнченнями збуджувальних нейронiв (див. рис. 2.9).
Активацiя аксо-аксонального синапса супроводжується деполяризацiєю пресинаптичної мембрани за типом катодичної депресiї, тобто натрiєвої iнактивацiї, до чого причетна кальцiєва проникнiсть. При надходженнi до такої деполяризованої дiлянки ПД затримується i тому у відповідному синапсi не видiляється медiатор. Але через iншi
синапси цей нейрон може збуджуватись, тобто пресинаптичне гальмування є фрагментарним. Фрагментарність бло- кування лише частки нейрона при пресинаптичному гальмуванні є однією з запорук його пластичності. Пресинаптич- не гальмування пригнічує в ЦНС несуттєві аферентні сигнали, тобто впливає на висхідні та низхідні провідникові шляхи спинного мозку, звільняючи від несуттєвої інформації, а в патологічному стані захищає мозок від зайвої аферентації, зокрема больової. Медiатором пресинаптичного гальмування є ГАМК.
Електричнi синапси
Міжнейронна передача збудження може відбуватися також електричним шляхом, тобто без участі медіаторів. Умовою для цього є щільний контакт між двома клітинами шириною до 9 нм. Отже, натрієвий струм від однієї з них може проходити через відчинені канали іншої мембрани. Тобто джерелом постси- наптичного струму другого нейрона є пресинаптична мембрана першого. Процес безмедіаторний; забезпечується виключно канальними білками (ліпідні мембрани для іонів непроникливі). Саме такі міжклітинні зв’язки названо нексусами (щілинними контактами). Вони розташовані строго один проти од- ного в мембранах двох нейронів – тобто на одній лінії; діаметром великі (до 1,5 нм у поперечнику), проникливі навіть для макромолекул масою до 1000. Складаються з субодиниць масою до 25000. Їх наявність звичайна для ЦНС як хребетних, так і безхребетних; властива групам синхронно функціонуючих клітин (зокрема, знайдені в мозочку між клітинами-зернами).
Бiльшiсть електричних синапсiв є збуджуючими. Але при певних морфологiчних характеристиках вони можуть бути гальмiвними. При двобічності проведення деякi з них мають випрямлюючий ефект, тобто проводять електричний струм значно краще вiд пресинаптичних структур до постсинаптичних, нiж у зво- ротному напрямку.
Дата добавления: 2015-12-11; просмотров: 1472;