РОЛЬ РАЗЛИЧНЫХ ЭЛЕМЕНТОВ НЕЙРОНА В ВОЗНИКНОВЕНИИ ВОЗБУЖДЕНИЯ
1. В возникновении ПД в нейронах в отличие от нервных и мышечных волокон (скелетные мышцы) принимают участие ионы Са2*, ток которых в клетку более медленный, чем ток ионов №+. В частности, в дендритах клеток Пуркинье мозжечка выявлены не только быстрые натриевые потенциалы, но и медленные кальциевые, вход ионов Са+ в пресинаптических окончаниях обеспечивает выброс медиатора. В телах некоторых нервных клеток ПД создается преимущественно за счет ионов Са2+, а в аксоне - главным образом за счет ионов Ка+.
2. Для возбуждения нейрона (возникновение ПД) необходимы поток афферентных импульсов и их взаимодействие. Это объясняется тем, что один пришедший к нейрону импульс обеспечивает выделение не более одного кванта медиатора (порция одного пузырька), что вызывает небольшой возбуждающий постсииаптиче-ский потенциал (ВПСП) - всего 0,05 мВ (миниатюрный ВПСП). Один пузырек содержит до десяти тысяч молекул медиатора, например ацетилхолина. Если учесть, что пороговый потенциал нейрона 5-10 мВ, то становится ясно, что для возбуждения нейрона требуется некоторое множество импульсов. Выброс медиатора из нервного окончания обеспечивает входящий в деполяризованную терминаль ток Са2+. Действует Са2+ с помощью белка кальмодулина. при этом 4 иона Са+ обеспечивают выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени в результате суммации ВПСП различных входов возникает деполяризация генераторного пункта, которая, достигнув критической величины, обеспечивает возникновение ПД нейрона-мишени. ВПСП возникает вследствие суммарного тока согласно электрохимическому градиенту в клетку и из клетки различных ионов через ионные каналы, функциональная активность которых определяется присутствием медиатора. Поступивший в пресинаптическое окончание Са2+ удаляется за его пределы с помощью Са-насоса. Действие выделившегося в синаптическую щель медиатора прекращается частично посредством обратного захвата его пресинаптическим окончанием, частично с помощью разрушения специальными ферментами. Норадреналин расщепляется моноаминоксидазой и катехолметил-трансферазой, ацетилхолин гидролизуется ацетилхолинтранс-феразой, имеющейся в синаптической щели и встроенной в постсинаптическую мембрану. Прекращение действия избытков медиатора на постсинаптическую мембрану предотвращает десенситизацию - снижение чувствительности постсипаптической мембраны к действующему медиатору.
Медиаторы, ферменты, белки, митохондрии транспортируются в пресинаптические окончания из тела клетки по аксону с помощью микротрубочек и микрофиламентов, тянущихся по всей длине аксона. Для этого транспорта необходимы ионы Са2* и энергия (АТФ непрерывно ресинтезируется в аксоне). Из синапса ретроградно транспортируются по аксону в тело клетки вещества, регулирующие в ней синтез белка.
3. Место возникновения ВПСП. Подавляющее большинство нейрональных синапсов находится на дендритах нейрона. Наиболее эффективно вызывают возбуждение нейрона синаптические контакты, расположенные на теле нейрона. Это связано с тем, что постсинаптические мембраны этих синапсов локализуются в непосредственной близости от места первичного возникновения ПД, в аксонном холмике. Близость соматических синапсов к аксонному холмику предполагает абсолютное участие их ВПСП в механизмах генерации ПД. В этой связи некоторые авторы предлагают называть их генераторными синапсами.
4. Место возникновения ПД. Генераторный пункт нейрона - ак-сонный холмик. Синапсы на нем отсутствуют. Отличительной особенностью аксонного холмика является высокая возбудимость, в 3-4 раза превосходящая возбудимость мембраны тела нейрона, что объясняется более высокой концентрацией Ка-каналов на аксонном холмике. ВПСП электротонически достигают аксонного холмика, обеспечивая здесь уменьшение мембранного потенциала до критического уровня. В этот момент возникает ПД. Возникший в аксонном холмике ПД, с одной стороны, ор-тодромно переходит на аксон, с другой - антидромно на тело нейрона (рис. 4.3). Как отмечалось, постоянная длины мембраны нейрона составляет 1 мм (расстояние, на котором ВПСП уменьшается на 37%), а диаметр тела нейрона в несколько десятков раз меньше, поэтому величина ВПСП, достигающая аксонного холмика, достаточна для возникновения на нем ПД.
При возбуждении нейронов потребление О2 возрастает в 2 раза, в цитоплазме уменьшается количество нуклеиновых кислот (иногда в 5 раз). Источником энергии является в основном глюкоза крови, собственные небольшие запасы гликогена достаточны лишь на 3-5 мин работы нейрона.
5. Роль дендритов в возникновении возбуждения до сих пор дискутируется. Дендритные синапсы удалены на значительное расстояние от генераторного пункта нейрона. По этой причине их ВПСП не могут вызвать там должную деполяризацию и обеспечить генерацию ПД. Считают, что синаптический аппарат денд-ритов проявляет себя при одновременном поступлении возбуждения к значительному числу дендритных синапсов. При этом суммарный дендритный ВПСП, изменяя мембранный потенциал генераторного пункта на подпороговом уровне, вызывает лишь модуляцию его возбудимости, делая возбудимость большей или меньшей в зависимости от временных и амплитудных характеристик колебаний мембранного потенциала генераторного пункта относительно величины критического уровня деполяризации. Данное обстоятельство, как выяснилось, может отразиться на выраженности ответной реащии нейрона при поступлении к нему в этот момент возбуждения через синапсы тела нейрона. Поэтому дендритные синапсы получили название модуляторных синапсов. >—
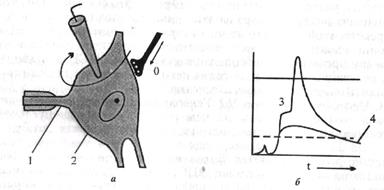
| Рис. 4.3. Регистрация (а) и временное течение ВПСП и ПД (б) в мотонейроне спинного мозга позвоночного при одиночной стимуляции заднекорешковых волокон. 1 - начальный сегмент аксона; 2 - сомадендритный комплекс; 3 - ПД; 4 – ВПСП 4.6. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ В ЦНС Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических |
синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие.
1. Иррадиация (дивергенция) возбуждения в ЦНС. Она объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 4.4, а).
Иррадиацию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное - энергичные движения всех конечностей и даже туловища. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
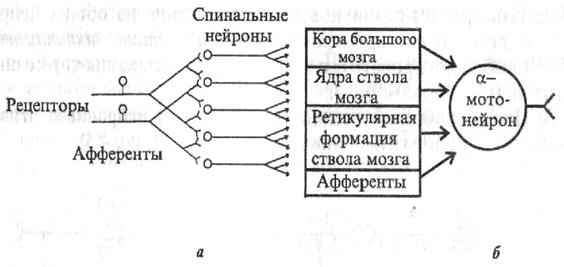
| Рис. 4.4. Дивергенция афферентных дорсальных корешков на спинальные нейроны, аксоны которых, в свою очередь, ветвятся, образуя многочисленные коллатерали (а), и конвергенция эфферентных путей от различных отделов ЦНС на а-мотонейрон спинного мозга (b) |
2. Конвергенция возбуждения (принцип общего конечного пути) - схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип шеррингтоновской воронки). Конвергенция возбуждения объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС может располагаться до 10 000 синапсов. Явление конвергенции возбуждения в ЦНС имеет широкое распространение. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Так, к одному и тому же спинальному мотонейрону подходят первичные афферентные волокна (рис. 4.4, 6). а также различные нисходящие пути многих вышележащих центров ствола мозга и других отделов ЦНС. Явление конвергенции весьма важно: оно обеспечивает, например, участие одного мотонейрона в нескольких различных реакциях. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг. На рис. 4.4, я показаны два афферентных волокна, каждое из которых отдает коллатерали к 4 нейронам таким образом, что 3 нейрона из общего их числа, равного 5, образуют связи с обоими афферентными волокнами. На каждом из этих 3 нейронов конвергируют два афферентных волокна. На один мотонейрон может конвергировать множество колла-тералей аксонов, до 10 000-20 000, поэтому генерация ПД в каждый момент зависит от общей суммы возбуждающих и тормозящих синаптических влияний. ПД возникают лишь в том случае, если преобладают возбуждающие влияния. Конвергенция может облегчать процесс возникновения возбуждения на общих нейронах в результате пространственной суммации подпороговых ВПСПлнбо блокировать его вследствие преобладания тормозных влияний (см. раздел 4.8).
3. Циркуляция возбуждения по замкнутым нейронным цепям. Она может продолжаться минуты и даже часы (рис. 4.5).
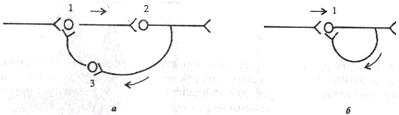
| Рис. 4.5. Циркуляция возбуждения в замкнутых нейронных цепях по Лоренто де-Но (а) и по И.С.Беритову (6). 1,2, 3 - возбуждающие нейроны Циркуляция возбуждения - одна из причин явления последействия, которое будет рассмотрено далее (см. раздел 4.7). Считают, |
что циркуляция возбуждения в замкнутых нейронных цепях - наиболее вероятный механизм феномена кратковременной памяти (см. раздел 6.6). Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом.
4. Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах. Распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении (см. раздел 4.3.3).
5. Замедленное распространение возбуждения в ЦНС по сравнению с его распространением по нервному волокну объясняется наличием на путях распространения возбуждения множества химических синапсов. Время проведения возбуждения через синапс затрачивается на выделение медиатора в синаптическую щель, распространение его до постсинаптической мембраны,
возникновение ВПСП и, наконец, ПД. Суммарная задержка передачи возбуждения в синапсе достигает примерно 2 мс. Чем больше синапсов в нейрональной цепочке, тем меньше общая скорость распространения по ней возбуждения. По латентному времени рефлекса, точнее по центральному времени рефлекса, можно ориентировочно рассчитать число нейронов той или иной рефлекторной дуги.
6. Распространение возбуждения в ЦНС легко блокируется определенными фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов.
Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию свойств нервных центров.
Дата добавления: 2015-10-22; просмотров: 1759;