СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Рассматриваемые ниже свойства нервных центров объясняются некоторыми особенностями распространения возбуждения в ЦНС, особыми свойствами химических синапсов и свойствами мембран нервных клеток. Основными свойствами нервных центров являются следующие.
А. Фоновая активность нервных центров (тонус) объясняется следующим:
• спонтанной активностью нейронов ЦНС;
• гуморальным влиянием циркулирующих в крови биологически активных веществ (метаболиты, гормоны, медиаторы и др.), влияющих на возбудимость нейронов;
• афферентной импульсацией от различных рефлексогенных зон;
• суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах;
• циркуляцией возбуждения в ЦНС.
Значение фоновой активности нервных центров заключается в обеспечении некоторого исходного уровня деятельного состояния центра и эффекторов. Этот уровень может возрастать или снижаться в зависимости от колебаний суммарной активности нейронов нервного центра-регулятора.
Б. Трансформация ритма возбуждения - это изменение числа импульсов, возникающих в нейронах центра на выходе, относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна как в сторону увеличения, так и в сторону уменьшения. Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию, способствуют иррадиация процесса возбуждения (см. раздел 4.6) и последействие. Уменьшение числа импульсов в нервном центре объясняется снижением его возбудимости за счет процессов пре- и постсинаптического торможения, а также избыточным потоком афферентных импульсов. При большом потоке афферентных влияний, когда уже все нейроны центра или нейронного пула возбуждены, дальнейшее увеличение афферентных входов не увеличивает число возбужденных нейронов.
В. Инерционность- сравнительно медленное возникновение возбуждения всего комплекса нейронов центра при поступлении к нему импульсов и медленное исчезновение возбуждения нейронов центра после прекращения входной импульсации. Инерционность центров связана с суммацией возбуждения и последействием.
I.Явление суммации возбуждения в ЦНС открыл И.М.Сеченов (1868) в опыте на лягушке: раздражение конечности лягушки слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождайся ответной реакцией - лягушка совершает прыжок. Различают временную (последовательную) суммацию и пространственную суммацию (рис. 4.6).
Временная сум мац и я. На рис. 4.6 слева показана схема для экспериментального тестирования эффектов, вызываемых в нейроне ритмической стимуляцией аксона. Запись вверху позволяет видеть, что если ВПСП быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временному ходу (несколько миллисекунд), достигая в конце концов порогового уровня. Временная суммация обусловлена тем, что ВПСП от предыдущего импульса еще продолжается, когда приходит следуюший импульс. Поэтому данный вид суммации называют также последовательной суммацией. Она играет важную физиологическую роль, потому что мнол-ге нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в нейронных объединениях нервных центров.
Пространственная суммация (см. рис. 4.6, б). Раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, тогда как при одновременной стимуляции обоих аксонов возникает ПД, что не может быть обеспечено одиночным ВПСП. Пространственная суммация связана с такой особенностью распространения возбуждения, как конвергенция.
2. Последействие - это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Причинами последействия являются:
• длительное существование ВПСП, если ВПСП полисинапти-ческий и высокоамплитудный; в этом случае при одном ВПСП возникает несколько ПД;
• многократные появления следовой деполяризации, что свойственно нейронам ЦНС; если следовая деполяризация достигает Е«р, то возникает ПД;
• циркуляция возбуждения по замкнутым нейронным цепям (см. раздел 4.6).
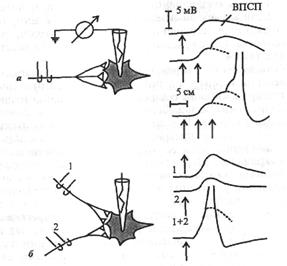
| Рис. 4.6. Суммация возбуждений в нейроне. а - временная суммация: один стимул (Т) и два стимула (ТТ) вызывают подпороговый ВПСП, три (ТТТ) последовательных стимула обеспечивают возникновение ПД; ((-пространственная суммация: раздельные одиночные раздражения вызывают подпороговые ВПСП, одновременные два раздражения вызывают ПД; 1, 2 - раздражающие электроды Первые две причины действуют недолго - десятки или сотни миллисекунд, третья причина - циркуляция возбуждения - может продолжаться минуты и даже часы. Таким образом, особенность распространения возбуждения (его циркуляция) обеспечивает другое явление в ЦНС - последействие. Последнее играет важнейшую роль в процессах обучения - кратковременной памяти. |
Г. Большая чувствительность ЦНС к изменениям внутренней среды:например, к изменению содержания глюкозы в крови, газового состава крови, температуры, к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостатку глюкозы и кислорода. При снижении содержания глюкозы в 2 раза ниже нормы могут возникнуть судороги. Тяжелые последствия для ЦНС вызывает недостаток-кислорода в крови. Прекращение кровотока всего на 10 с приводит к очевидным нарушениям функций мозга: человек теряет сознание. Если кровоток прекращается на 8-12 мин, то возникают необратимые нарушения деятельности мозга; погибают многие нейроны, в первую очередь корковые, что ведет к тяжелым последствиям.
Д. Утомляемость нервных центровпродемонстрировал Н. Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения большеберцового (n. tibialis) и малоберцового (n. peroneus) нервов. В этом случае ритмическое раздражение одного нерва вызывает ритмическое сокращение мышцы, приводящее к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги (рис. 4.7).
При этом развивается постсииаптическая депрессия(привыкание, габитуация) - ослабление реакции центра на раздражения (афферентные импульсы), выражающееся в снижении постсинаптических потенциалов во время длительного раздражения или после него. Это ослабление объясняется расходованием медиатора, накоплением метаболитов, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
Е. Пластичность нервных центров- способность нервных элементов к перестройке функциональных свойств. Основные проявления этого свойства следующие: посттетаническая потен-циация и депрессия, доминанта, образование временных связей, а в патологических случаях - частичная компенсация нарушенных функций.
1. Посттетаническая потенциация (синаптическое облегчение) - это улучшение проведения в синапсах после короткого раздражения афферентных путей. Кратковременная активация увеличивает
амплитуду постсинаптических потенциалов. Облегчение наблюдается и во время раздражения (вначале); в этом случае феномен называют тетанической потенциацией. Степень выраженности облегчения возрастает с увеличением частоты импульсов; облегчение максимально, когда импульсы поступают с интервалом в несколько миллисекунд.
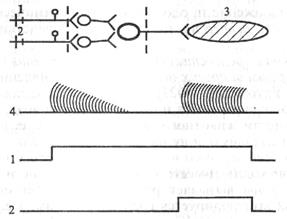
| Рис. 4.7. Схема опыта Н.Е.Введенского, иллюстрирующего локализацию утомления в рефлекторной дуге. I - раздражение большеберцового нерва; 2 – раздражение малоберцового нерва; 3 - полусухожильная мышца лягушки; 4 – кривая сокращения полусухожилыюй мышцы Длительность посттетанической потенциации зависит от свойств синапса и характера раздражения. После одиночных стимулов она выражена слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. По-видимому, главной причиной возникновения синаптического облегчения является накопление Са2+ в пресинапти- |
ческих окончаниях, поскольку Са2+, который входит в нервное окончание во время ПД, накапливается там, так как ионная помпа не успевает выводить его из нервного окончания. Соответственно увеличивается высвобождение медиатора при возникновении каждого импульса в нервном окончании, возрастает ВПСП. Кроме того, при частом использовании синапсов ускоряются синтез медиатора и мобилизация пузырьков медиатора; напротив, при редком использовании синапсов синтез медиаторов уменьшается - важнейшее свойство ЦНС. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах. Значение синаптического облегчения, по-видимому, заключается в том, что оно создает предпосылки улучшения процессов переработки информации на нейронах нервных центров, что крайне важно, например, для обучения в ходе выработки условных рефлексов. Повторное возникновение явлений облегчения в нервном центре может вызвать переход центра из обычного состояния в доминантное.
2. Если раздражение продолжается, то в химических синапсах может наступить депрессия, по-видимому, вследствие истощения медиатора.
3. Доминанта - господствующий очаг возбуждения в ЦПС, подчиняющий себе функции других нервных центров. Явление доминанты открыл А.А.Ухтомский (1923) в опытах с раздражением двигательных зон большого мозга и наблюдением за возникающим сгибанием конечности животного. Как выяснилось, если раздражать корковую двигательную зону на фоне избыточного повышения возбудимости другого нервного центра, то обычного сгибания конечности не происходит. Вместо сгибания конечности раздражение двигательной зоны вызывает реакцию тех эффекторов, деятельность которых контролируется господствующим, т.е. доминирующим в данный момент в ЦНС, нервным центром.
В эксперименте доминанту можно получить многократной посылкой афферентных импульсов к определенному центру, гуморальными на него влияниями. Роль гормонов в образовании доминантного очага возбуждения демонстрирует опыт на лягушке: весной у самца раздражение любого участка кожи вызывает не защитный рефлекс, а усиление обнимательного рефлекса. В условиях натурального поведения доминантное состояние нервных центров может быть вызвано метаболическими причинами.
Доминантный очагвозбуждения обладает рядом особых свойств,главными из которых являются следующие: инерционность, стойкость, повышенная возбудимость, способность «притягивать» к себе иррадиирующие по ЦНС возбуждения, способность оказывать угнетающие влияния на центрыконкуренты и другие нервные центры.
Значениедоминантного очага возбуждения в ЦНС заключается в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов. Например, на базе доминантного состояния центра голода реализуется пищедобывательное поведение; на базе доминантного состояния центра жажды запускается поведение, направленное на поиск воды. Успешное завершение данных поведенческих актов в итоге устраняет физиологические причины доминантного состояния центров голода и жажды. Доминанта играет важную роль в координационной деятельности ЦНС (см. раздел 4.9), в запоминании и переработке информации.
4. Компенсация нарушенных функций после повреждения того или иного центра - результат проявления пластичности ЦНС. Хорошо известны клинические наблюдения, когда у больных после кровоизлияний в вещество мозга повреждались центры регуляции мышечного тонуса и акта ходьбы. Тем не менее, со временем отмечалось, что парализованная конечность постепенно начинает вовлекаться в двигательную активность, при этом нормализуется тонус ее мышц. Нарушенная двигательная функция частично, а иногда и полностью восстанавливается за счет большей активности сохранившихся нейронов и вовлечения в эту функцию других - «рассеянных» - нейронов в коре большого мозга с подобными функциями. Этому способствуют регулярные пассивные и активные движения.
ТОРМОЖЕНИЕ В ЦНС
Торможение- это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение вторично относительно процесса возбуждения, так как всегда возникает как следствие возбуждения.
Торможение в ЦНС открыл И.М.Сеченов(1863), В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И.М.Сеченова послужило толчком для дальнейших исследований торможения в ЦНС. В частности, Ф.Гольц (1870) обнаружил проявления торможения у спинальной лягушки. Он также исследовал латентное время рефлекса. Оказалось, что механическое раздражение кончиков пальцев одной конечности лягушки существенно удлиняет латентный период сгибательного рефлекса другой конечности при погружении ее в раствор кислоты. Наличие специальных тормозных структур в продолговатом мозге доказал Х.Мегун (1944). В опытах на кошках при изучении разгибательного рефлекса он установил, что раздражение медиальной части ретикулярной формации продолговатого мозга тормозит рефлекторную активность спинного мозга.
Тонкий анализ тормозных явлений в ЦНС позволил выделить две разновидности торможения: 1) постсинаптическое торможение и 2) пресинаптическое торможение. 4.8.1. Постсинаптическое торможение
А. Этот вид торможения открыл Д.Экклс (1952) при регистрации потенциалов мотонейронов спинного мозга у кошки во время сокращения и расслабления мышцы в ходе реализации соответствующих рефлекторных актов. Оказалось, что при рефлекторном расслаблении мышц на мотонейронах регистрируется гиперполяризационный постсшшптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был назван тормозным постсинстттеским потенциалом (ТПСП; рис. 4.8).
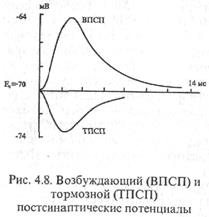
| У кошки ТПСП регистрируется на 0,5 мс позже, чем ВПСП, что объясняется наличием на пути проведения возбуждения, запускающего ТПСП, одного дополнительного синапса. Амплитуда ТПСП 1-5 мВ. Он способен суммироваться, более мощный афферентный залп вызывает возрастание амплитуды ТПСП Б. Механизм постсинаптического торможения. Возбудимость клетки от ТПСП (гиперполяризационного постсинаптического потенциала) уменьшается, потому что увеличивается пороговый потенциал (V), так как Екр (критический уровень деполяризации КУД) остается на прежнем уровне, а мембранный потенциал (Ео) возрастает. ТПСП возникает под влиянием аминокислоты глицина. Глицин выделяется особыми тормозными клетками в синапсах, образуемых этими |
клетками на мембране нейрона-мишени. Действуя на постсинаптическую мембрану, глицин увеличивает ее проницаемость для С1- и в меньшей степени для К+. Оба иона движутся согласно концентрационным градиентам и вопреки электрическим градиентам: С1- в клетку, К+ из клетки, в результате развивается гиперполяризация. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической суммации ТПСП и ВПСП, в связи с чем в зоне ак-сонного холмика не происходит выведения мембранного потенциала на критический уровень. Как известно, вышедший из клетки К+ транспортируется обратно в клетку Nа/К-помпой, что и поддерживает градиент его концентрации. По-видимому, и С1 транспортируется в этом случае из клетки специальной хлорной помпой, хотя обычно отмечают, что градиент концентрации СЬ поддерживается отрицательным электрическим зарядом внутри клетки - С1 выталкивается отрицательным электрическим зарядом. Возникновение ТПСП обеспечивается входом С1- в клетку. Это доказывает, что концентрационный градиент действует сильнее противоположно направленного электрического градиента. Отсутствие С1--помпы привело бы к исчезновению химического (концентрационного) градиента для С1 и нарушению процесса торможения. Но этого не происходит, что подтверждает подобное заключение. Кроме спинного мозга, гли-цинорецепторы обнаружены на нейронах ствола мозга. Как выяснилось, ТПСП могут возникать вследствие уменьшения проницаемости мембраны для Ка+, что также сопровождается гиперполяризацией клеточной мембраны, особенно если проницаемость для К+ и С1 сохраняется прежней. Такого рода ТПСП были зарегистрированы в нейронах симпатических ганглиев.
В. Разновидности постсинаптического торможения. Обычно выделяют возвратное, латеральное, параллельное и прямое (реципрокное) Постсинаптическое торможение. Имеются и другие варианты классификаций. Некоторые авторы называют только два торможения - возвратное и прямое, последнее трактуется по-разному. В реальной действительности вариантов торможения больше, они определяются множеством связей различных нейронов, в частности их кол.чатералей.
1. Возвратное постсшшптическое торможение — это такое торможение, когда тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. В этом случае развивающееся торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Типичным примером возвратного постсинаптического торможения является торможение в мотонейронах спинного мозга. Как показано на рис. 4.9, 2, мотонейроны посылают коЯлатерали к тормозным вставочным нейронам, аксоны которых в свою очередь образуют синапсы на тех же мотонейронах, которые возбуждают тормозную клетку Реншоу. Такая тормозная цепь называется торможением Реншоу - в честь ученого, который ее открыл, а тормозные вставочные нейроны в этой цепи - клетками Реншоу. Это торможение в центрах сгибателей и разгибателей обеспечивает, например, поочередное сокращение и расслабление скелетной мышцы, что необходимо при ходьбе и беге. Сама клетка Реншоу возбуждается под влиянием ацетилхолина с помощью М-холинорецептора.
2. Подобную роль может играть и параллельное торможение, когда возбуждение блокирует само себя за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возратом импульсов к нейрону, который активировался этим же возбуждением (см. рис. 4.9, 1).
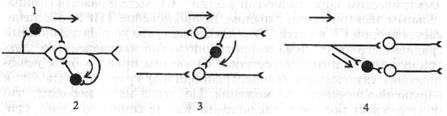
| Рис. 4.9. Разновидности постсинаптического торможения. 1 - параллельное; 2 - возвратное; 3 - латеральное; 4 - прямое; нейроны; О—< возбуждающие, 0—< тормозные 3. Латеральное постсинаптическое торможение иллюстри- |
рует рис. 4.9
3. Тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в этих соседних клетках развивается очень глубокое торможение. Такого типа торможение называется латеральным потому, что образующаяся зона торможения находится «сбоку» по отношению к возбужденному нейрону и инициируется им. Латеральное торможение играет особенно важную роль в афферентных системах. Латеральное торможение может образовать тормозную зону, которая окружает возбуждающие нейроны.
4. Примером прямого торможения может служить реципрокное торможение. Оно вызывает угнетение центра-антагониста. Например, при раздражении кожных рецепторов возникает защитный сгибательный рефлекс - центр сгибания возбужден, а центр разгибания заторможен. В этом случае возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку Реншоу - к центру мышцы-антагониста - разгибателю, что предотвращает ее сокращение (см. рис. 4.9, 4). Если бы одновременно возбуждались центры мышц - сгибателей и разгибателей, сгибание конечности в суставе было бы невозможным.
Дата добавления: 2015-10-22; просмотров: 1306;