Строение половых клеток
У высших животных при половом размножении, сопровождающемся оплодотворением, осуществляется не только передача наследственной информации, но большую роль играет эндокринная регуляция процессов гаметогенеза и сложные преобразования, связанные с дроблением зиготы, развитием, дифференцировкой органов и тканей. До оплодотворения происходит образование гамет. Гаметы у млекопитающих формируются в половых железах в результате гаметогенеза. По сравнению с другими клетками функция гамет уникальна. Они обеспечивают передачу наследственной информации между особями разных поколений, чем сохраняют жизнь во времени. Был период в биологии, когда половые и соматические клетки противопоставляли друг другу, наделяя лишь первые всей полнотой свойств жизни, проносимых ими через поколения. В настоящее время экспериментально доказана возможность развития полноценного организма на основе наследственной информации ядра дифференцированной соматической клетки, например кишечного эпителия. В 1990-х годах группе английских ученых, использовавших принципиально аналогичный подход, удалось вырастить (клонировать) овцу Долли. Это был пример успешного решения задачи получения взрослой особи на основе генетического материала соматической клетки у млекопитающих. Напомним, что клоны представлены генетически идентичными клетками, образовавшимися путем последовательных митотических делений исходно одной клетки-родоначальницы и ее потомков. Вытекающая из этого техническая возможность клонирования людей порождает серьезные морально-этические проблемы, так как допускает преднамеренное «тиражирование» избранного человека с присущим только ему набором физических психо-эмоциональных свойств и даже интеллектуального потенциала. В настоящее время общественностью, в том числе и научной, признается недопустимым клонирование людей. Вместе с тем, использование технологии клонирования для получения достаточного числа клеток определенного типа, генетически неотличимых от клеток данного организма и, следовательно, иммунологически с ним совместимых, рассматривается как перспективное направление в области трансплантации тканей.Гаметы представляют собой одно из многих направлений дифференцировки клеток многоклеточного организма. Они образуют «клеточную линию», специализированную к выполнению репродуктивной функции. Предполагают, что клетки указанной линии возникают из бластомеров, имеющих на вегетативном полюсе цитоплазму особого рода — так называемую зародышевую плазму, богатую РНК.
Гаметы - высокодифференцированные клетки, содержащие наследственную информацию, необходимую для развития организма. По ряду признаков женские и мужские гаметы отличаются друг от друга, что связано с различными функциями яйцеклетки и сперматозоида в процессе размножения.
Яйцеклетки неподвижны, имеют ядро, цитоплазму, питательный материал (желток). По содержанию желтка клетки могут быть алецитальными, изолецитальными, телолецитальными, центролецитальными. Алецитальные яйцеклетки содержат очень мало желтка. Они характерны для плацентарных млекопитающих и для человека. Изолецитальныеяйцеклетки мелкие, с небольшим количеством равномерно распределенного желтка. Такие яйцеклетки характерны для хордовых, двустворчатых и брюхоногих моллюсков. Телолецитальные яйцеклетки - могут быть с умеренным или с большим содержанием желтка. Яйцеклетки рыб и земноводных имеют умеренное содержание желтка. Он находится на одном из полюсов, названном вегетативным. Полюс, не содержащий желтка, назван анимальным. Пресмыкающиеся, птицы имеют телолецитальные яйцеклетки с очень большим содержанием желтка. На анимальном полюсе находится зародышевый диск с ядром и цитоплазмой без желтка. На вегетативном полюсе желтка много. У центролецитальных яйцеклеток желток находится вокруг ядра. По периферии клетки расположена свободная от желтка цитоплазма. Эти яйцеклетки характерны для членистоногих.
Полярность яйцеклеток намечается еще на стадии накопления желтка в ооцитах во время их быстрого роста и закрепляется при выделении полярных телец именно на анимальном полюсе. Внутреннюю разнокачественность участков яйца называют ооплазматической сегрегацией. После оплодотворения (у асцидий уже через 5 мин) в еще не дробящемся яйце происходит закономерное перераспределение цитоплазмы. В дальнейшем цитоплазма разного состава также закономерно распределяется по клеткам тканей разных зачатков. По-видимому, на ранних стадиях способность бластомеров развиваться в определенном направлении зависит от наследования ими веществ, концентрирующихся в разных участках цитоплазмы яйцеклетки.
Кортикальный слой — это поверхностный слой цитоплазмы яйца вместе с плазматической мембраной. В нем находятся микрофиламенты и кортикальные гранулы. Кортикальный слой играет важную роль в кортикальной реакции при оплодотворении.
Яйцеклетки имеют оболочки, которые выполняют защитную функцию, обеспечивают требуемый уровень обмена веществ, препятствуют проникновению в яйцеклетку более одного спермия, способствуют внедрению (имплантации) зародыша в стенку матки у плацентарных животных, поддерживают форму зародыша.
Лицевые оболочки. Различают первичную оболочку, образуемую самой яйцевой клеткой, вторичную оболочку, являющуюся продуктом деятельности фолликулярных клеток, и третичные оболочки, которыми яйцо окружается во время прохождения по яйцеводу.
Первичная оболочка, иногда называемая желточной, имеется у яйцеклеток всех животных. У позвоночных, в том числе млекопитающих и человека, первичная оболочка входит в состав плотной оболочки, образуя ее внутреннюю часть. Внешняя часть плотной оболочки продуцируется фолликулярными клетками и является вторичной оболочкой. Плотная оболочка изнутри пронизана микроворсинками яйцеклетки, а снаружи — микроворсинками фолликулярных клеток, отчего при большом увеличении она выглядит исчерченной и названа лучистой оболочкой (corona radiata). За свои оптические свойства у млекопитающих она получила название блестящей оболочки (zona pellucida). Таким образом, эта оболочка совмещает в себе первичную и вторичную оболочки. Третичные оболочки хорошо развиты у хрящевых рыб и амфибий, но особенную сложность они приобретают у наземных позвоночных — пресмыкающихся, птиц и низших млекопитающих. Образуясь из секретов желез яйцевода, эти оболочки не имеют клеточного строения. У всех позвоночных они выполняют функции защиты зародыша от механических повреждений и действия вредных биотических факторов, таких, как бактериальные, грибковые и протозойные. Кроме того, у наземных позвоночных появляются принципиально новые функции запаса воды и питательных веществ для обеспечения нужд зародыша. У пресмыкающихся скорлуповая оболочка действует как насос, забирая воду из почвы и воздуха. У птиц запас воды находится в белковой оболочке. Поглощение и испарение воды регулируется порами в скорлуповой оболочке. Скорлупа содержит множество минеральных солей, необходимых для развития скелета зародыша.
Мужские половые клетки - сперматозоиды - очень мелкие и способны двигаться. Сперматозоиды млекопитающих имеют головку, шейку и хвост. Головка содержит ядро и немного цитоплазмы. В шейке есть центриоли и митохондрии. От шейки отрастает хвост, представляющий собой жгутик, необходимый для передвижения. Сперматозоид имеет аппарат движения в виде жгутика. В семенной жидкости мужская гамета человека развивает скорость до 5 см/ч. Поясним приводимую цифру следующим примером. Если учесть соотношение преодолеваемого расстояния и длины движущегося объекта, то при названной скорости сперматозоид человека перемещается в 1,5 раза быстрее, чем пловец олимпийского ранга. Яйцеклетка, лишенная аппарата активного движения, преодолевает расстояние до полости матки, равное примерно 10 см, за 4—7 сут. Сперматозоиды некоторых видов животных имеют акросомный аппарат, выбрасывающий при контакте с яйцеклеткой длинную нить. Он обеспечивает проникновение ядра мужской гаметы в цитоплазму яйцеклетки путем растворения особыми ферментами ее оболочек. Описаны и другие приспособления, способствующие оплодотворению.
В сравнении с другими линиями соматических клеток (эпителиальные, нервные, мышечные) гаметы характеризуются рядом отличий:
1) Гаплоидный наборхромосом в ядрах, что обеспечивает воспроизведение в зиготе типичного для организмов данного вида диплоидного числа хромосом. Действительно, оплодотворение сперматозоидом яйцеклетки, ядра которых содержат по 23 хромосомы, обусловливает формирование зиготы с 46 хромосомами, что типично для соматических клеток человека.
2) Необычное значение ядерно-цитоплазматического отношения. У яйцеклеток оно снижено благодаря увеличенному объему цитоплазмы, в которой размещен питательный материал (желток) для развития зародыша. У сперматозоидов благодаря малому количеству цитоплазмы ядерно-цитоплазматическое отношение высокое. Это находится в соответствии с главной функциональной задачей мужской гаметы — транспортировкой наследственного материала к яйцеклетке.
3) Низкий уровень обменных процессов, близкий к состоянию анабиоза. Мужские гаметы не вступают в митотический цикл.У яйцеклеток эта способность восстанавливается при оплодотворении или действии фактора, активирующего партеногенез.
Гаметогенез– процесс образования половых клеток: яйцеклеток (оогенез) и сперматозоидов (сперматогенез).
Клетки зачаткового эпителия половых желез делятся последовательно митозом и мейозом. В результате этих делений образуются мужские гаметы (сперматозоиды) и женские гаметы (яйцеклетки). Они имеют гаплоидный набор хромосом и содержат наследственную информацию, необходимую для развития организма.
Гаметогенез подразделяется на ряд стадий.
Встадии размножения диплоидные клетки, из которых образуются гаметы, называют сперматогониямии оогониями. Эти клетки осуществляют серию последовательных митотических делений, в результате чего их количество существенно возрастает. Сперматогонии размножаются на протяжении всего периода половой зрелости мужской особи. Сперматогонии развиваются из первичных половых клеток, мигрирующих в семенники на ранней стадии эмбрионального развития. Наружный слой клеток стенок канальцев семенника содержит диплоидный набор хромосом. Клетки делятся митозом. Их число увеличивается. Образуются незрелые половые клетки -сперматогонии. Они имеют округлую форму и крупное ядро.
Клетки зачаткового эпителия делятся митозом и образуются оогонии. Размножение оогоний приурочено главным образом к периоду эмбриогенеза. У человека в женском организме этот процесс наиболее интенсивно протекает в яичниках между 2-м и 5-м месяцами внутриутробного развития. К 7-му месяцу большая часть ооцитов входит в профазу I мейоза. Так как способом размножения клеток-предшественниц женских и мужских гамет является митоз, то оогонии и сперматогонии, как и все соматические клетки, характеризуются диплоидностью. В ходе митотического цикла их хромосомы имеют либо однонитчатую (после митоза и до завершения синтетического периода интерфазы), либо двунитчатую (постсинтетический период, профаза и метафаза митоза) структуру в зависимости от количества биспиралей ДНК. Если в одинарном, гаплоидном наборе число хромосом обозначить как n, а количество ДНК — как с, то генетическая формула клеток в стадии размножения соответствует 2n2сдо S-периода и 2n4c после него.
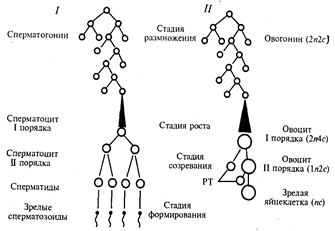
Схема гаметогенеза:
1 — сперматогенез, 2 — овогенез, n — количество хромосомных наборов,
с — количество ДНК, РТ — редукционные тельца
На стадии роста происходит увеличение клеточных размеров и превращение мужских и женских половых клеток в сперматоцитыи ооциты I порядка, причем последние достигают больших размеров, чем первые. Сперматогонии перемещаются в зону роста, расположенную ближе к просвету канальца. Клетки увеличиваются в размерах и называются сперматоцитами 1-го порядка.
Ооциты I порядка до полового созревания остаются на стадии профазы I мейотического деления. Ооциты 1-го порядка на этой стадии могут оставаться очень долго (десятки лет). С наступлением половой зрелости каждый месяц один из ооцитов 1-го порядка увеличивается в размерах, окружается фолликулярными клетками, обеспечивающими питание. Одна часть накапливаемых веществ представляет собой питательный материал (желток в ооцитах), другая — связана с последующими делениями. Важным событием этого периода является репликация ДНК при сохранении неизменным числа хромосом. Последние приобретают двунитчатую структуру, а генетическая формула сперматоцитов и ооцитов I порядка приобретает вид 2n4с.
Основными событиями стадии созревания являются два последовательных деления: редукционное и эквационное, которые вместе составляют мейоз. После первого деления образуются сперматоциты и ооциты II порядка (формула n2с), а после второго — сперматидыизрелая яйцеклетка (nс).С наступлением половой зрелости сперматоциты постепенно претерпевают мейотические деления. В этом периоде происходят два мейотических деления. Каждый сперматоцит 1-го порядка в результате 1-го мейотического деления образует два сперматоцита 2-го порядка с гаплоидным набором хромосом. После второго мейотического деления возникают еще по две сперматиды. Это овальные клетки небольших размеров.
Под влиянием гормонов ооцит 1-го порядка заканчивает 1-е мейотическое деление и образуется один ооцит 2-го порядка и полярное (редукционное) тельце. 2-е деление мейоза идет до стадии метафазы. На этой стадии ооцит 2-го порядка выходит из яичника в брюшную полость, а оттуда попадает в яйцевод. Но дальнейшее созревание не произойдет, пока ооцит не соединится со сперматозоидом. В яйцеводах ооцит 2-го порядка заканчивает 2-е деление мейоза и образует оотиду- крупную клетку, и второе полярное тельце. Таким образом, из одной оогонии образуется одна оотида и три полярных тельца. Редукционные тельца разрушаются. Процесс образования половых клеток регулируется гормонами. Если оплодотворения не произойдет, ооцит 2-го порядка погибнет и будет выведен из организма.
Процесс сперматогенеза завершается стадией формирования, или спермиогенеза. сперматиды перемещаются ближе к просвету канальца. Из сперматид формируются сперматозоиды определенного строения, способные к передвижению. Хвосты сперматозоидов направлены в просвет канальца. Таким образом, из одного сперматогония формируются 4 зрелых сперматозоида, которые выходят в просвет семенного канальца. Сперматогенез регулируется гормонами. Ядра сперматид уплотняются вследствие сверхспирализации хромосом, которые становятся функционально инертными. Пластинчатый комплекс перемещается к одному из полюсов ядра, образуя акросомный аппарат, играющий большую роль в оплодотворении. Центриоли занимают место у противоположного полюса ядра, причем от одной из них отрастает жгутик, у основания которого в виде спирального чехлика концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, так что головка зрелого сперматозоида практически ее лишена. У яйцеклеток период формирования отсутствует. Продолжительность развития зрелых сперматозоидов из родоначальных клеток - сперматогоний - составляет около 72 дней. Сформированный сперматозоид имеет длину около 70 мкм. В ядре сперматозоида человека содержится 23 хромосомы, одна из которых является половой (X и Y), остальные - аутосомами. Среди сперматозоидов 50% содержат Х-хромосому и 50% - Y-хромосому. Показано, что масса Х-хромосомы больше массы Y-хромосомы, поэтому сперматозоиды, содержащие Х-хромосому, менее подвижны, чем содержащие Y-хромосому.
Сперматогенез отличается от оогенеза в нескольких отношениях:
1) После полового созревания в мейоз непрерывно вступают новые клетки;
2) Из каждой приступившей к мейозу клетки образуется не одна, а четыре зрелые гаметы;
3) Зрелые спермии формируются после завершения мейоза в ходе сложного процесса клеточной дифференцировки.
Оплодотворение — это процесс слияния сперматозоида и яйцеклетки, сопровождающийся объединением геномов отцовского и материнского организмов и завершающийся образованием зиготы, которая представляет собой начальный этап развития нового организма. У человека оплодотворение яйцеклетки происходит в яйцеводах. В зиготе возрастает ядерно-плазменное соотношение. Резко усиливаются обменные процессы. Зигота способна к дальнейшему развитию. Сущность оплодотворения состоит во внесении сперматозоидом отцовских хромосом. Сперматозоид оказывает стимулирующее влияние, вызывающее начало развития яйцеклетки
Оплодотворению предшествует осеменение, обеспечивающее встречу мужских и женских гамет. Осеменение может быть наружным и внутренним. Наружное осеменение характерно для животных, обитающих в воде (рыбы, амфибии). Яйцеклетки и сперматозоиды выделяются в воду, и там происходит их слияние. Внутреннее осеменение характерно для животных, обитающих на суше. Сперматозоиды во время полового акта вводятся в половые пути самки. Встреча гамет осуществляется в верхних отделах яйцеводов. Продолжительность жизни и способность у яиц и сперматозоидов к оплодотворению ограничены. У большинства беспозвоночных и водных позвоночных животных яйца должны быть сразу оплодотворены после овуляции. Яйца большинства млекопитающих сохраняют способность к оплодотворению на протяжении 24 часов, у человека 12-24 часа после овуляции. Сперматозоиды сохраняют способность к оплодотворению в половых путях самки в течение нескольких часов. Продолжительность жизни сперматозоидов и сохранение ими оплодотворяющей способности зависит от внешних факторов (освещенности, температуры, концентрации углекислого газа, водородных ионов) в среде, в которой находятся гаметы. Оплодотворение может произойти лишь при определенной концентрации сперматозоидов в семенной жидкости. Обычно в 1 мл семенной жидкости мужчины содержится около 350 млн. сперматозоидов. Искусственное осеменение применяют для выведения ценных пород животных. Селекционеры разработали способы консервации спермы элитных производителей с сохранением сперматозоидами оплодотворяющей способности. При некоторых условиях сперма может быть заморожена и сохраняться до 6 лет. Искусственное осеменение и оплодотворение в пробирке с последующей трансплантацией позволило женщинам, которые ранее не могли иметь детей, испытать счастье материнства. За время, прошедшее после рождения первого ребенка «из пробирки» в 1978 г., оплодотворение in vitro позволило лучше изучить влияние возраста и других факторов на способность к зачатию. При этом методе яйцеклетка человека оплодотворяется в лаборатории, затем ее переносят в матку, где завершается развитие плода. Встреча гамет осуществляется независимо от нормального полового процесса. Это помогает наблюдать явления, которые обычно трудно изучить, поскольку они протекают в организме женщины. Знание механизмов оплодотворения и первых этапов развития яиц млекопитающих может помочь в лечении бесплодия. После осеменения происходит оплодотворение. У млекопитающих яйцеклетка сохраняет способность к оплодотворению в течение суток, сперматозоиды - несколько часов. Для успешного оплодотворения нужна определённая концентрация сперматозоидов в семенной жидкости (для человека это 200 млн. на 1 мл), вероятность выделения гаметами особых веществ - гино- и андрогамонов, повышает вероятность контакта их. У человека объём эякулята в норме составляет около 3 мл; в нём содержится в среднем 350 млн. сперматозоидов. Для обеспечения оплодотворения общее количество сперматозоидов в сперме должно быть не менее 150 млн., а концентрация их в 1 мл - не менее 60 млн. В половых путях женщины после копуляции их число уменьшается по направлению к дистальному концу маточной трубы. Благодаря высокой подвижности сперматозоиды при оптимальных условиях могут через 30 мин - 1 час достигать полости матки, а через 1,5-2 часа находиться в дистальной части маточной трубы, где происходит встреча с яйцеклеткой и оплодотворение. Сперматозоиды сохраняют оплодотворяющую способность до 2 суток.
Процесс оплодотворения складывается из трех последовательных фаз: а) сближения гамет; б) активации яйцеклетки; в) слияния гамет, или сингамии.
1. Сближение сперматозоида с яйцеклеткой обеспечивается совокупностью неспецифических факторов, повышающих вероятность их встречи и взаимодействия. К ним относят скоординированность наступления готовности к оплодотворению у самца и самки, поведение самцов и самок, обеспечивающее совокупление и осеменение, избыточную продукцию сперматозоидов, крупные размеры яйцеклетки, а также вырабатываемые яйцеклетками и сперматозоидами химические вещества, способствующие сближению и взаимодействию половых клеток. Эти вещества, называемые гамонами (гормоны гамет), с одной стороны, активируют движение сперматозоидов, а с другой — их склеивание. В особой структуре сперматозоида — акросоме — локализуются протеолитические ферменты. У млекопитающих большое значение имеет пребывание сперматозоидов в половых путях самки, в результате чего мужские половые клетки приобретают оплодотворяющую способность (капацитация), т.е. способность к акросомной реакции.
В момент контакта сперматозоида с оболочкой яйцеклетки происходит акросомная реакция, во время которой под действием протеолитических ферментов акросомы (гиалуронидаза и др.) яйцевые оболочки растворяются. Далее плазматические мембраны яйцеклетки и сперматозоида сливаются и через образующийся вследствие этого цитоплазматический мостик цитоплазмы обеих гамет объединяются. Затем в цитоплазму яйца переходят ядро и центриоль сперматозоида, а мембрана сперматозоида встраивается в мембрану яйцеклетки. Хвостовая часть сперматозоида у большинства животных тоже входит в яйцо, но потом отделяется и рассасывается, не играя какой-либо роли в дальнейшем развитии.
2. В результате контакта сперматозоида с яйцеклеткой происходит ее активация. Она заключается в сложных структурных и физико-химических изменениях. Яйцеклетка женщины (как и млекопитающих животных) вторично изолецитального типа, содержит небольшое количество желточных зёрен, более или менее равномерно расположенных в ооплазме. Свой резерв питательных веществ яйцеклетка человека обычно расходует в течение 12 - 24 ч после овуляции, а затем погибает, если не будет оплодотворена. Оптимальные условия для взаимодействия сперматозоидов с яйцеклеткой обычно создаются в пределах 12 ч после овуляции. При осеменении многочисленные спермин приближаются к яйцеклетке и вступают в контакт с ее оболочкой. Яйцеклетка начинает совершать вращательные движения вокруг своей оси со скоростью 4 вращения в минуту. Эти движения обусловлены влиянием биения жгутиков сперматозоидов и продолжаются около 12 ч. Благодаря тому, что участок мембраны сперматозоида проницаем для ионов натрия, последние начинают поступать внутрь яйца, изменяя мембранный потенциал клетки. Затем в виде волны, распространяющейся из точки соприкосновения гамет, происходит увеличение содержания ионов кальция, вслед за чем также волной растворяются кортикальные гранулы. Выделяемые при этом специфические ферменты способствуют отслойке желточной оболочки; она затвердевает, это оболочка оплодотворения. Все описанные процессы представляют собой так называемую кортикальную реакцию. Одним из значений кортикальной реакции является предотвращение полиспермии, т.е. проникновения в яйцеклетку более одного сперматозоида. У млекопитающих кортикальная реакция не вызывает образования оболочки оплодотворения, но суть ее та же.
У таких животных, как морской еж, костистые рыбы и земноводные, все изменения цитоплазмы сопровождаются видимыми морфологическими перестройками. Эти явления получили название расслоения или сегрегации плазмы.
Активация яйцеклетки завершается началом синтеза белка на трансляционном уровне, поскольку мРНК, тРНК, рибосомы и энергия были запасены еще в оогенезе. Активация яйцеклетки может начаться и протекать до конца без ядра сперматозоида и без ядра яйцеклетки, что доказано опытами по энуклеации зиготы.
3. Яйцеклетка в момент встречи со сперматозоидом обычно находится на одной из стадий мейоза, заблокированной с помощью специфического фактора. У большинства позвоночных этот блок осуществляется на стадии метафазы II; у многих беспозвоночных, а также у трех видов млекопитающих (лошади, собаки и лисицы) блок происходит на стадии диакинеза. В большинстве случаев блок мейоза снимается после активации яйцеклетки вследствие оплодотворения. В то время как в яйцеклетке завершается мейоз, ядро сперматозоида, проникшее в нее, видоизменяется. Оно принимает вид интерфазного, а затем профазного ядра. За это время удваивается ДНК и мужской пронуклеус получает количество наследственного материала, соответствующего, т.е. содержит гаплоидный набор редуплицированных хромосом.
Ядро яйцеклетки, закончившее мейоз, превращается в женский пронуклеус, также приобретая n2с. Оба пронуклеуса проделывают сложные перемещения, затем сближаются и сливаются (синкарион), образуя общую метафазную пластинку. Это, собственно, и есть момент окончательного слияния гамет — сингамия. Первое митотическое деление зиготы приводит к образованию двух клеток зародыша (бластомеров) с набором хромосом 2n2c в каждом.
Дата добавления: 2015-10-09; просмотров: 3071;