НР1-залежна система репресії
Білок НР1 (Heterochromatin Protein 1) містить два структурні домени: хромодомен (модуль, що впізнає метильовані лізинові залишки) і так званий хромошедоу (chromoshadow) – домен, який має спорідненість до певних специфічних HD і гістонметилтрансферази (HMT), а також здатен взаємодіяти з іншою молекулою НР1.
Прикладом залучення НР1 до утворення гетерохроматину є центромери хромосом (рис. 25). Деацетилювання Lys9 і Lys14 гістону Н3 сприяє зв’язуванню специфічної НМТ, яка здійснює метилювання Lys9 гістону Н3. Цей метильований Lys (MeLys9) упізнається хромодоменом НР1. Завдяки хромошедоу домену НР1 рекрутує HD, яка підтримує деацетильований статус суміжних нуклеосом, і ту саму НМТ, що здійснює метилювання Lys9 у складі суміжних нуклеосом: знов виникає лавиноподібний процес, що самопідтримується та розповсюджується на сусідні ділянки. Взаємодія між білками НР1 (деталі якої не з’ясовані) забезпечує додаткову компактизацію фібрили. Природа обмежувача, який не дозволяє гетерохроматину розповсюджуватися поза центромерну зону, не з’ясована, але гранична ділянка містить свій специфічний маркер – метильований Lys4 гістону Н3 (і деметильований Lys9).
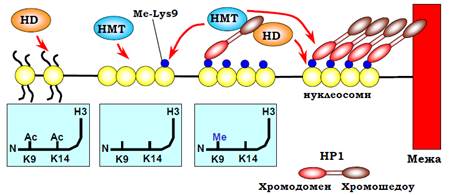
Рис. 25. Розповсюдження та самопідтримання гетерохроматинового стану в центромерах. На вставках: стан модифікацій двох ключових залишків у N-кінцевому хвості гістону Н3.
При реплікації білки тимчасово знімаються з ДНК у точці реплікаці. Реплікація ДНК здійснюється нерівномірно – гетерохроматин реплікується в останню чергу. За точкою реплікації гістони батьківського хроматину (які несуть на собі гетерохроматинові маркери) повертаються на дочірні молекули ДНК разом із гістонами, синтезованими de novo, НР1 також повертається на той самий локус, де він був присутній, і відновлює патерни модифікацій гістонів і компактний (репресований) стан гетерохроматинової ділянки, тобто, гетерохроматиновий стан даного локусу відтворюється в дочірніх клітинах. При цьому НР1 має спорідненість до білків ламіни, що зумовлює розташування гетерохроматину в периферичних зонах клітинного ядра.
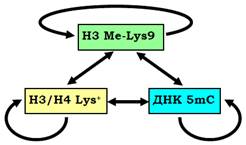
Рис. 26. Взаємний вплив деацетилювання гістонів,метилювання Lys9 гістону Н3 і метилювання ДНК у гетерохроматині.
Подібна система репресії за участю НР1 широко використовується в інших ділянках гетерохроматину, а також для гарантованого блокування генів в еухроматинових зонах. Метилювання Lys9 гістону Н3 та деацетильований стан лізинів гістонів Н3/Н4 є найхарактернішою ознакою таких ділянок. Як описано вище, обидві модифікації самопідримуються та підтримують одна одну через опосередковану дію НР1. При цьому важливу роль у забезпеченні репресованого стану відіграє метилювання цитозинів ДНК – інша ковалентна модифікація, яка також відновлюється при клітинному поділі й замикає своєрідне коло репресії (рис. 26).
Дата добавления: 2015-09-11; просмотров: 1046;