Транскрипційні фактори
Відповіддю на необхідність регулювати окремо активність великої кількості генів лімітованим набором факторів транскрипції є принцип модульності будови еукаріотичнихпромоторів. Цей принцип ілюструє рис. 10: три регуляторні (наприклад, проксимальні) елементи послідовності мають спорідненість до трьох транскрипційних факторів, із трьох пар такої взаємодії можна скласти шість комбінацій. Насправді таких пар значно більше, і кількість можливих комбінацій – практично нескінченно велика. Кожен промотор може мати свій власний набір модулів, який відрізняє його від інших промоторів, і, відповідно, власний набір досить великої кількості транскрипційних факторів, потрібних для активації гена.

Рис. 10. Модульний принцип взаємодії транскрипційних факторів із промоторами.
Із принципом модульності тісно пов’язаний принцип кооперативності взаємодії транскрипційних факторів із цис-елементами, що знаходяться поряд. Кожен з ТФ зазвичай має порівняно невисоку спорідненість до відповідних елементів послідовності. Але якщо два цис-елементи розташовані поруч, і два ТФ здатні взаємодіяти між собою, спорідненість кожного до них підвищується (рис. 11): стабільність комплексу значно зростає (експоненціально) завдяки сумі невеликих знижень вільної енергії при взаємодії кожного окремого ТФ. Приклад на рис. 11 показує лише один елементарний акт кооперативної взаємодії кількох ТФ із промотором.

Рис. 11. Приклад кооперативної взаємодії двох транскрипційних факторів із ДНК (1A02).
Крім того, більшість факторів транскрипції мають у своїй структурі принаймні два домени: той, що взаємодіє з ДНК, і так званий активаційний (AD, Activation Domain), використовується для взаємодії з іншими білками (рис. 10). Активацйні домени зв’язують білкові кофактори (коактиватори), у результаті на дистальних і проксимальних елементах промотора формується складний мультибілковий комплекс – енхансосома (enhanceosome). До складу енхансосом часто входять також гістонацетилтрансферази та фактори ремоделювання хроматину (див. нижче).
Різні елементи енхансосоми діють синергічно, підвищуючи загальну стабільність комплексу. З іншого боку, відсутність кількох елементів може зумовити дестабілізацію та розпад енхансосоми, деспорідненість до ДНК кожного окремого елемента є невисокою.
Це забезпечує динамізм активації: енхансосома не є фіксованою, а збирається / розбирається в певні моменти.
Активаційні домени ТФ і коактиватори мають, у свою чергу, спорідненість до медіатору та базальних факторів транскрипції. Результатом такої взаємодії є ефективне збирання преініціаторного комплексу на базальному промоторі (рис. 12). Слід зауважити, що певні компоненти мультибілкових комплексів, які збираються на промоторах, можуть, навпаки, блокувати ініціацію транскрипції – тоді їх називають репресорами та корепресорами.
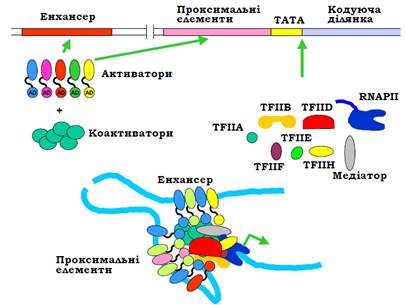
Рис. 12. Схема збирання комплексу активації транскрипції.
Дата добавления: 2015-09-11; просмотров: 1525;